Abstract
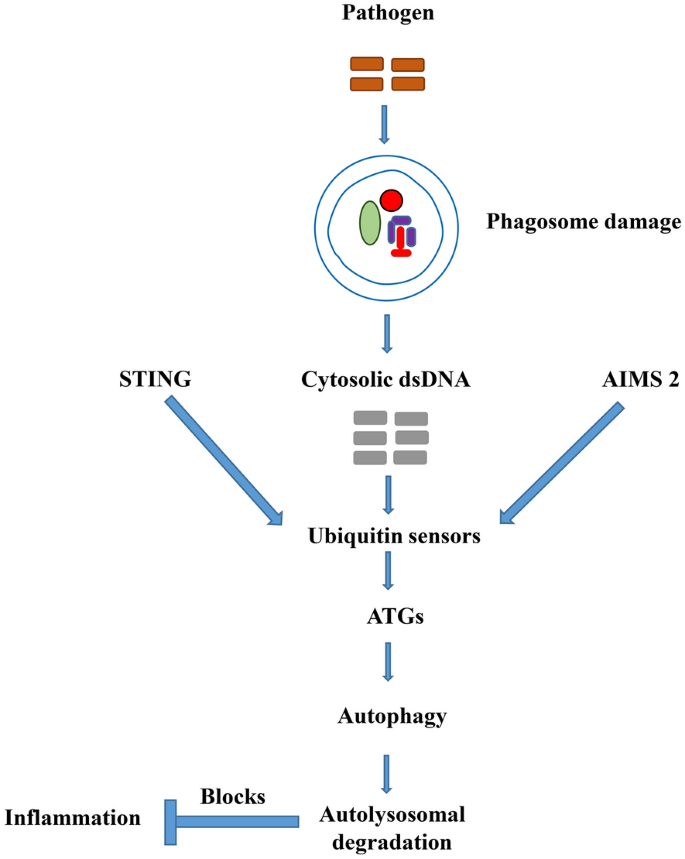
BACKGROUND: Autophagy has a crucial role in the defense against parasites. The interplay existing between host autophagy and parasites has varied outcomes due to the kind of host cell and microorganism. The presence of autophagic compartments disrupt a significant number of pathogens and are further cleared by xenophagy in an autolysosome. Another section of pathogens have the capacity to outwit the autophagic pathway to their own advantage. RESULT: To comprehend the interaction between pathogens and the host cells, it is significant to distinguish between starvation-induced autophagy and other autophagic pathways. Subversion of host autophagy by parasites is likely due to differences in cellular pathways from those of 'classical' autophagy and that they are controlled by parasites in a peculiar way. In xenophagy clearance at the intracellular level, the pathogens are first ubiquitinated before autophagy receptors acknowledgement, followed by labeling with light chain 3 (LC3) protein. The LC3 in LC3-associated phagocytosis (LAP) is added directly into vacuole membrane and functions regardless of the ULK, an initiation complex. The activation of the ULK complex composed of ATG13, FIP200 and ATG101causes the initiation of host autophagic response. Again, the recognition of PAMPs by conserved PRRs marks the first line of defense against pathogens, involving Toll-like receptors (TLRs). These all important immune-related receptors have been reported recently to regulate autophagy. CONCLUSION: In this review, we sum up recent advances in autophagy to acknowledge and understand the interplay between host and parasites, focusing on target proteins for the design of therapeutic drugs. The target host proteins on the initiation of the ULK complex and PRRs-mediated recognition of PAMPs may provide strong potential for the design of therapeutic drugs against parasitic infections.