1. 领域背景与文献引入
文献英文标题:Evolutionary history and functional implications of protein domains and their combinations in eukaryotes;发表期刊:Genome Biology;影响因子:未明确提供;研究领域:真核生物蛋白质组学与进化生物学
蛋白质结构域作为蛋白质的基本功能与结构单位,是蛋白质进化的核心研究对象,自1990年代SCOP、CATH等蛋白质结构分类数据库建立以来,结构域的进化研究逐渐成为进化生物学的热点方向。随着真核生物基因组测序的推进,研究者发现多细胞真核生物的蛋白质结构域组合复杂度显著提升,推测其与细胞黏附、信号传导等复杂多细胞功能的进化密切相关。现有研究已证实结构域组合遵循无标度网络特性,功能相关的结构域易发生融合形成多结构域蛋白,但不同真核生物类群中结构域组合的进化机制差异尚未得到系统解析,尤其是动物类群与其他真核生物在构建复杂多细胞系统时的进化策略是否存在本质区别,仍是领域内未解决的核心问题。
针对这一研究空白,本研究通过整合47种真核生物的基因组数据,系统解析了不同类群中蛋白质结构域及其组合的特异性进化模式,旨在阐明真核生物结构域组合的进化历史与功能意义,为理解复杂多细胞系统的进化机制提供新的视角。
2. 文献综述解析
作者在综述部分以真核生物的进化类群(动物、真菌、植物、原生生物)和结构域的进化年龄为核心分类维度,系统梳理了领域内关于蛋白质结构域组合的研究进展,明确了现有研究的结论、优势与局限性,并通过对比凸显了本研究的创新价值。
现有研究的关键结论包括,蛋白质结构域组合的伙伴数量遵循幂律分布,古老结构域通常具有更高的连接度,功能相关的结构域倾向于通过基因融合形成多结构域蛋白,从而拓展蛋白质的功能多样性。技术方法上,现有研究多依赖Pfam等标准化结构域数据库进行注释,结合比较基因组学分析结构域的进化模式,具有注释统一、可重复性高的优势。但现有研究也存在明显局限性,多数研究集中在真核生物、细菌、古菌三界的跨域比较,缺乏针对真核生物内部不同类群的系统分析,尤其是动物类群中结构域组合的快速进化机制未被深入解析,无法明确不同真核生物类群在构建复杂多细胞系统时的进化策略差异。
本研究的创新点在于首次基于47种真核生物的大规模基因组数据,针对真核生物内部不同类群进行结构域组合的特异性分析,揭示了动物类群通过高连接度的特异性结构域实现功能多样化的独特进化策略,而其他真核生物类群则依赖古老结构域的重复利用,这一发现填补了领域内关于真核生物类群特异性结构域进化机制的研究空白,为理解复杂多细胞系统的进化提供了新的理论依据。
3. 研究思路总结与详细解析
本研究的核心目标是解析真核生物蛋白质结构域组合的进化规律,阐明其与功能多样化的关系;核心科学问题是不同真核生物类群中结构域组合的进化机制差异,尤其是动物类群的特异性进化策略;技术路线遵循“结构域注释→特异性鉴定→连接度分析→网络进化分析→结论”的闭环逻辑,通过多层面的生物信息学分析系统揭示结构域组合的进化模式。
3.1 蛋白质结构域注释与组合定义
实验目的:建立标准化的真核生物蛋白质结构域注释体系,明确结构域组合的定义标准,为后续的进化分析奠定基础。
方法细节:采用Pfam数据库(14.0版)的结构域注释体系,对47种真核生物(涵盖脊椎动物、昆虫、线虫、真菌、植物、原生生物)的蛋白质组进行结构域分配,使用HMMER软件进行结构域比对,阈值设置为E值<10^-3;定义结构域组合为蛋白质序列中连续相邻的两个结构域,且区分结构域的顺序(如A+B与B+A被视为不同的组合)。
结果解读:共从47种真核生物的蛋白质组中注释得到4315个Pfam结构域,鉴定出6977种独特的结构域组合;多细胞动物(尤其是脊椎动物)的蛋白质平均结构域数量显著高于其他类群(文献未明确提供具体数值,基于图表趋势推测);真菌的结构域覆盖度(结构域覆盖的氨基酸残基占总序列的比例)略高于其他类群,平均约为20%-30%(文献未明确提供具体数值,基于图表趋势推测)。
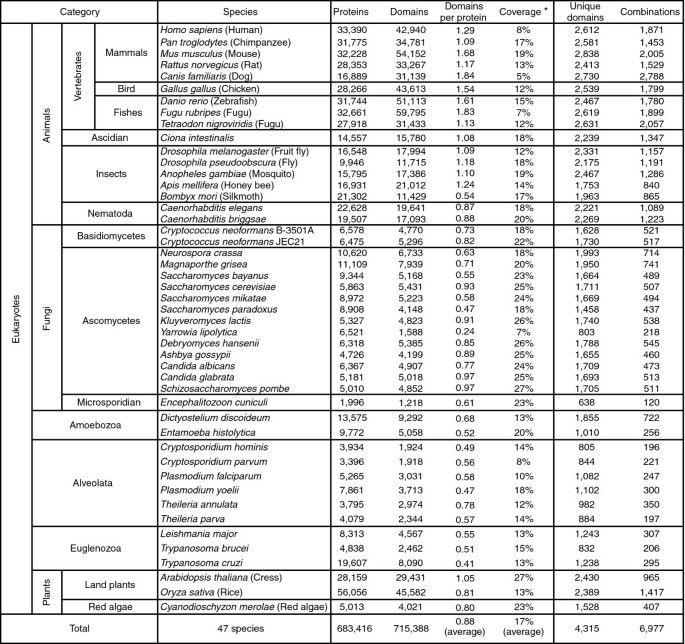
产品关联:文献未提及具体实验产品,领域常规使用Pfam数据库、HMMER软件进行蛋白质结构域注释,使用生物信息学分析平台(如KEGG)进行基因组数据整合。
3.2 类群特异性结构域及组合的鉴定
实验目的:鉴定不同真核生物类群的特异性结构域及其组合,明确其进化起源与类群分布特征。
方法细节:基于简约性进化模型,重建结构域在真核生物进化过程中的获得与丢失事件,区分真核生物特异性结构域(仅存在于真核生物)与类群特异性结构域(仅存在于某一真核生物类群,如动物、后口动物);对于结构域组合,同样通过简约性模型判断其是否为类群特异性起源,排除独立进化产生的相同组合。
结果解读:共鉴定得到3104个真核生物特异性结构域,其中动物特异性结构域组合达875种,后口动物(脊椎动物加尾索动物)特异性组合达610种;动物特异性结构域组合主要涉及酪氨酸磷酸化信号通路,包括蛋白激酶结构域、SH2结构域、蛋白酪氨酸磷酸酶结构域等;后口动物特异性组合主要涉及补体与凝血级联反应,包括胰蛋白酶样丝氨酸蛋白酶结构域、血管性血友病因子A型结构域等。
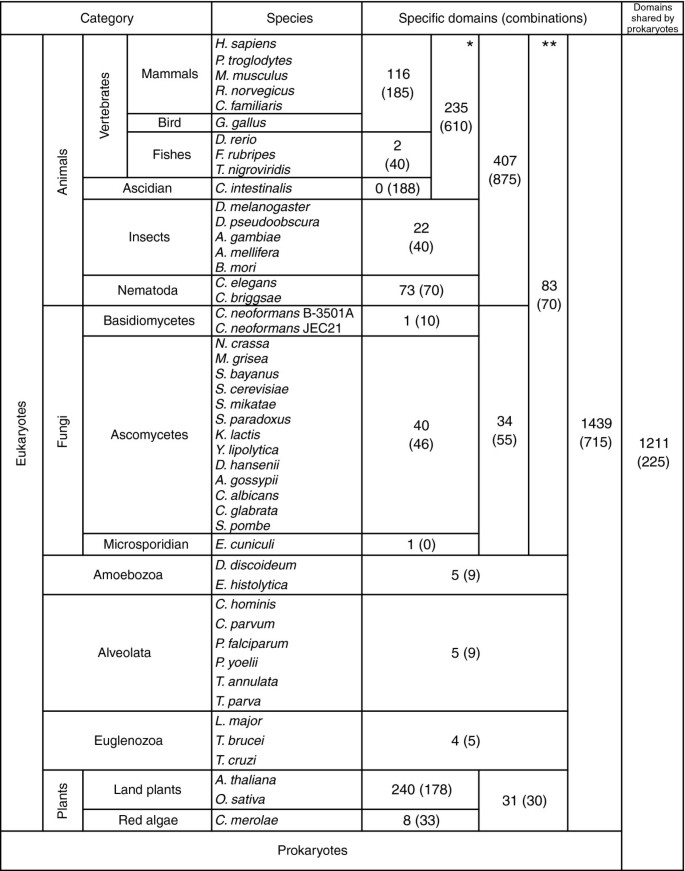
产品关联:文献未提及具体实验产品,领域常规使用进化分析软件(如MEGA、PhyML)进行简约性进化模型重建,使用自定义脚本进行类群特异性分析。
3.3 结构域连接度与类群特异性分析
实验目的:分析不同类群中结构域的连接度(即结构域的组合伙伴数量)差异,揭示结构域组合的进化驱动因素。
方法细节:统计不同类群特异性结构域的平均连接度,绘制连接度随进化层级的变化趋势图;对比动物类群(后口动物、无脊椎动物)与非动物类群(植物、真菌、原生生物)的结构域连接度模式。
结果解读:动物类群中,类群特异性结构域的平均连接度显著高于古老结构域(如从真核生物与原核生物共同祖先继承的结构域);而非动物类群中,古老结构域的平均连接度更高,呈现出连接度随结构域进化年龄增加而升高的趋势;动物特异性结构域(如蛋白激酶结构域)的连接度最高,是形成大量特异性组合的核心驱动因素。
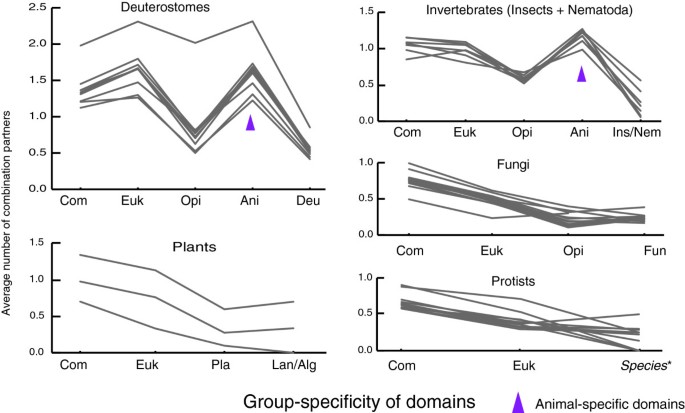
产品关联:文献未提及具体实验产品,领域常规使用生物信息学工具(如Cytoscape、NetworkX)进行网络连接度分析,使用统计软件(如R)进行趋势图绘制。
3.4 结构域组合网络的全局进化分析
实验目的:从全局网络层面解析真核生物结构域组合的进化模式,对比动物与真菌的网络进化差异。
方法细节:构建结构域组合网络(节点为结构域,边为结构域组合),分析网络的无标度特性(幂律分布);追踪人类与酿酒酵母谱系中网络幂律指数γ的进化变化,对比实际网络与联合网络(包含所有祖先结构域组合,不考虑丢失)的差异。
结果解读:人类谱系中,幂律指数γ在动物与真菌分化后快速下降,表明结构域组合网络呈现加速生长的特性;酿酒酵母谱系中,γ值逐渐上升,而联合网络的γ值显著下降,提示基因丢失是其网络进化的主要因素;动物类群的网络加速生长依赖于高连接度的特异性结构域,这是动物类群实现功能多样化的关键进化机制。
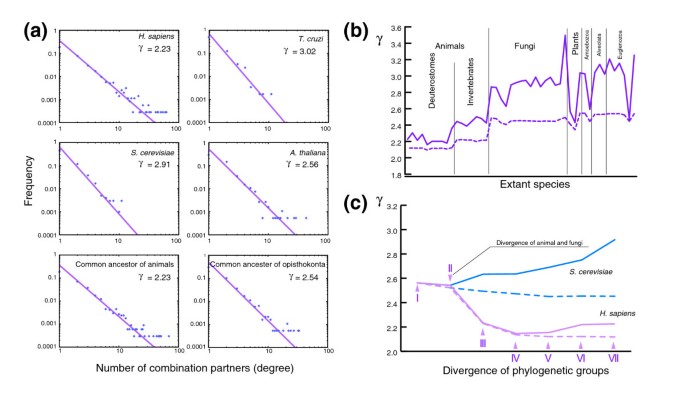
产品关联:文献未提及具体实验产品,领域常规使用网络分析工具(如NetworkX)进行幂律分布拟合,使用进化分析软件进行谱系追踪。
4. Biomarker研究及发现成果
本文中的Biomarker为真核生物不同类群的特异性蛋白质结构域及其组合,可作为类群进化的分子标志物,其筛选与验证基于大规模基因组数据的进化分析,揭示了不同真核生物类群的独特进化策略。
Biomarker定位:本研究中鉴定的类群特异性结构域及其组合,属于进化层面的分子标志物,筛选逻辑为:基于47种真核生物的基因组数据,通过简约性进化模型鉴定仅存在于某一类群的结构域及组合,验证其在对应类群中的分布特异性,排除跨类群分布的结构域及组合。
研究过程详述:这些Biomarker的来源为47种真核生物的蛋白质组数据,验证方法为进化起源分析与类群分布统计;动物特异性结构域组合仅存在于动物类群,涉及酪氨酸磷酸化信号通路,是动物类群信号传导功能进化的关键;后口动物特异性组合仅存在于后口动物,涉及补体与凝血级联反应,是后口动物先天免疫系统进化的核心;文献未明确提供特异性与敏感性的量化数据。
核心成果提炼:这些类群特异性结构域组合可作为真核生物类群进化的可靠分子标志物,揭示了动物类群通过高连接度的特异性结构域实现功能多样化的独特进化策略,而其他真核生物类群则依赖古老结构域的重复利用;该发现为理解真核生物复杂多细胞系统的进化机制提供了新的视角,为后续研究不同类群的功能进化奠定了基础,文献未明确提供统计学显著性数据。