Abstract
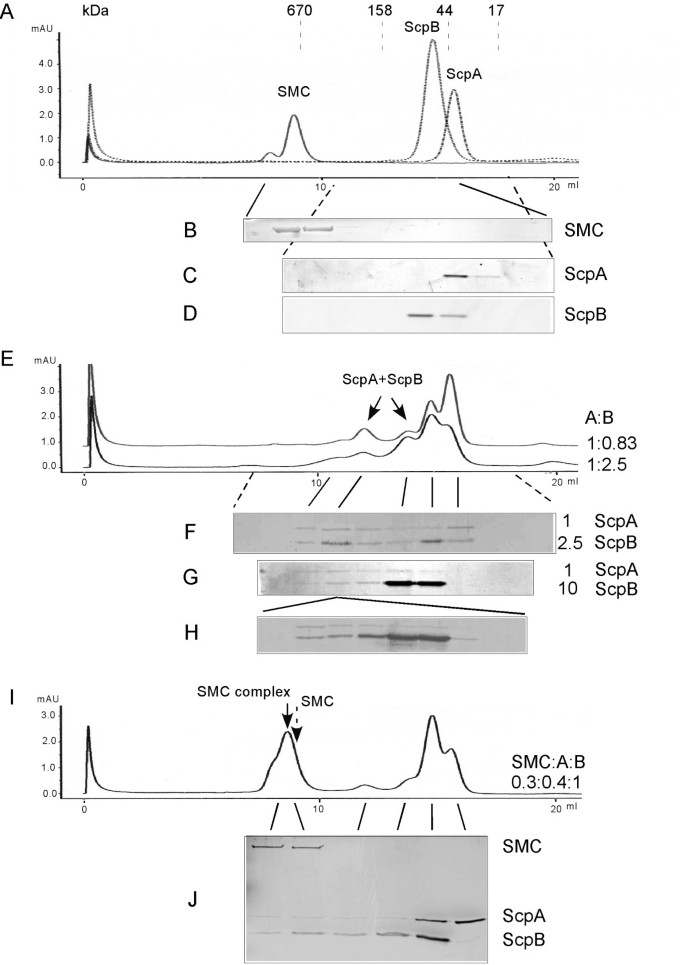
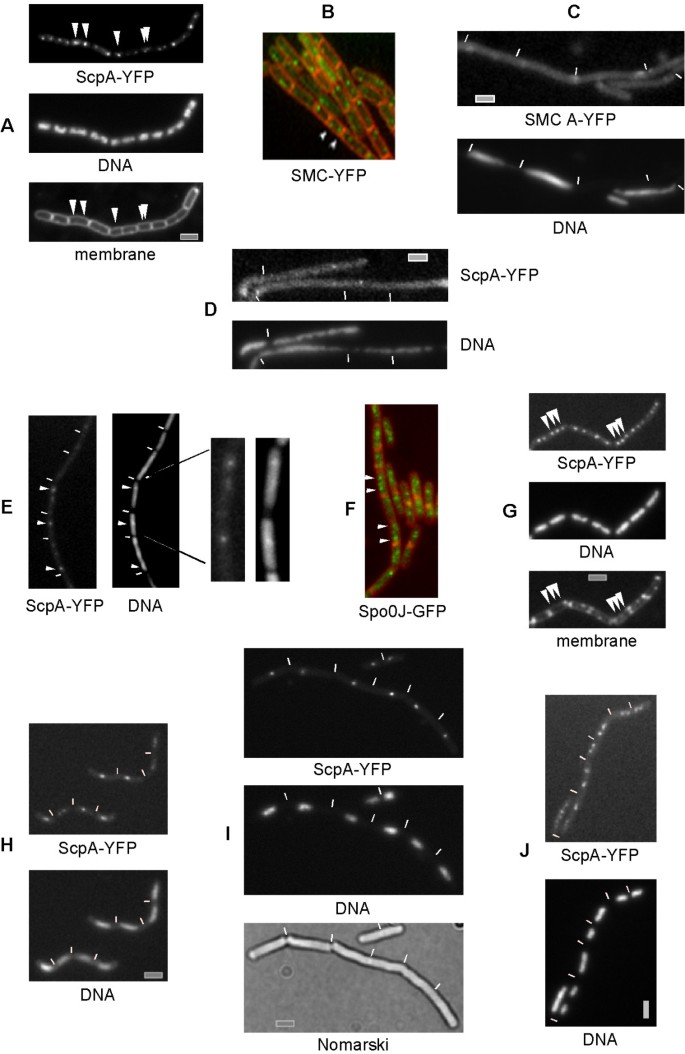
BACKGROUND: SMC proteins are key components of several protein complexes that perform vital tasks in different chromosome dynamics. Bacterial SMC forms a complex with ScpA and ScpB that is essential for chromosome arrangement and segregation. The complex localizes to discrete centres on the nucleoids that during most of the time of the cell cycle localize in a bipolar manner. The complex binds to DNA and condenses DNA in an as yet unknown manner. RESULTS: We show that in vitro, ScpA and ScpB form different complexes with each other, among which the level of the putative 2 ScpA/4 ScpB complex showed a pronounced decrease in level upon addition of SMC protein. Different mutations of the ATPase-binding pocket of SMC reduced, but did not abolish interaction of mutant SMC with ScpA and ScpB. The loss of SMC ATPase activity led to a loss of function in vivo, and abolished proper localization of the SMC complex. The formation of bipolar SMC centres was also lost after repression of gyrase activity, and was abnormal during inhibition of replication, resulting in single central clusters. Resumption of replication quickly re-established bipolar SMC centres, showing that proper localization depends on ongoing replication. We also found that the SMC protein is subject to induced proteolysis, most strikingly as cells enter stationary phase, which is partly achieved by ClpX and LonA proteases. Atomic force microscopy revealed the existence of high order rosette-like SMC structures in vitro, which might explain the formation of the SMC centres in vivo. CONCLUSION: Our data suggest that a ScpA/ScpB sub-complex is directly recruited into the SMC complex. This process does not require SMC ATPase activity, which, however, appears to facilitate loading of ScpA and ScpB. Thus, the activity of SMC could be regulated through binding and release of ScpA and ScpB, which has been shown to affect SMC ATPase activity. The proper bipolar localization of the SMC complex depends on a variety of physiological aspects: ongoing replication, ATPase activity and chromosome supercoiling. Because the cellular concentration of SMC protein is also regulated at the posttranscriptional level, the activity of SMC is apparently regulated at multiple levels.