Background
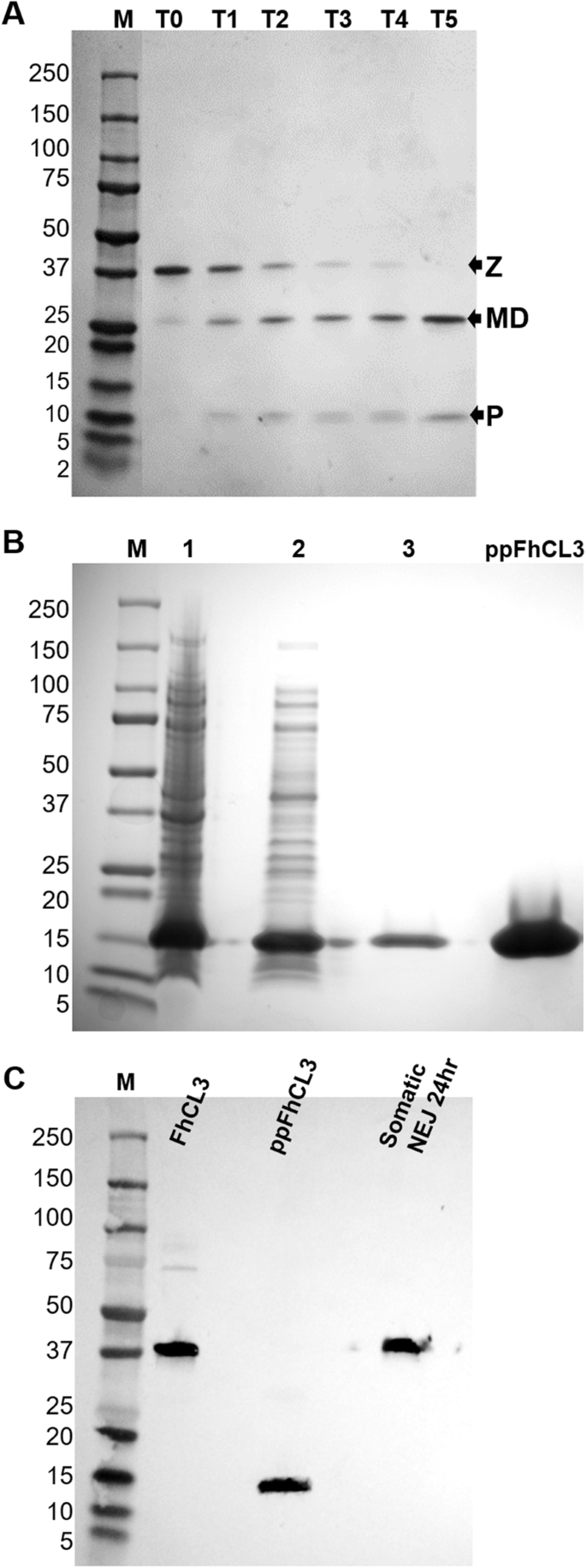
The zoonotic worm parasite Fasciola hepatica secretes an abundance of cathepsin L peptidases that are associated with virulence, invasiveness, feeding and migration. The peptidases are produced as inactive zymogens that activate at low pH by autocatalytic removal of their N-terminal pro-domain or propeptide. Propeptides bind to their cognate enzyme with high specificity. Little is known, however, about the mechanism by which the propeptide of FhCL3, a cathepsin L peptidase secreted by the infective newly excysted juveniles (NEJs), regulates the inhibition and activation of the mature enzyme before it is secreted into host tissues.
Conclusions
The FhCL3 peptidase involved in host invasion by F. hepatica is produced as a zymogen in the NEJs gut. Regulation of its activation involves specific binding sites within the propeptide that are interdependent and act as a "clamp-like" mechanism of inhibition. These interactions are disrupted by the low pH of the NEJs gut to initiate autocatalytic activation. Our enzyme kinetics data demonstrates high potency and selectivity of the ppFhCL3 for its cognate FhCL3 enzyme, information that could be utilised to design inhibitors of parasite cathepsin L peptidases.
Results
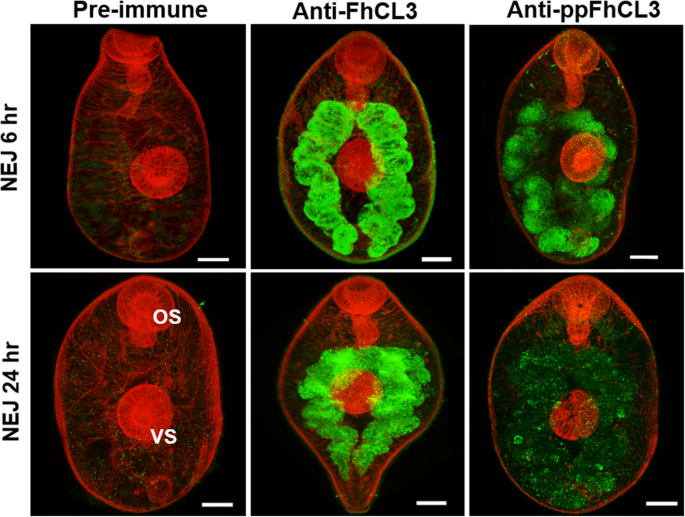
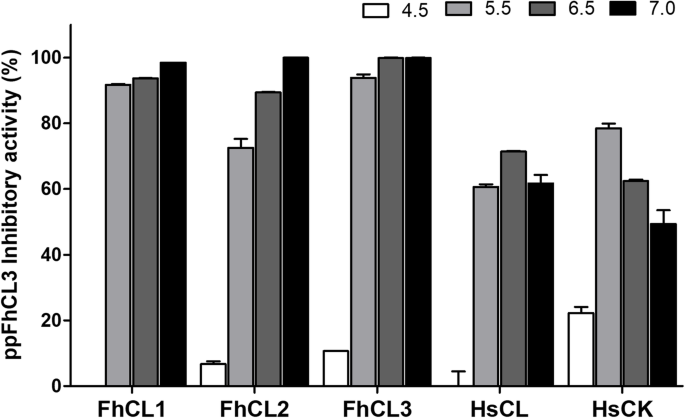
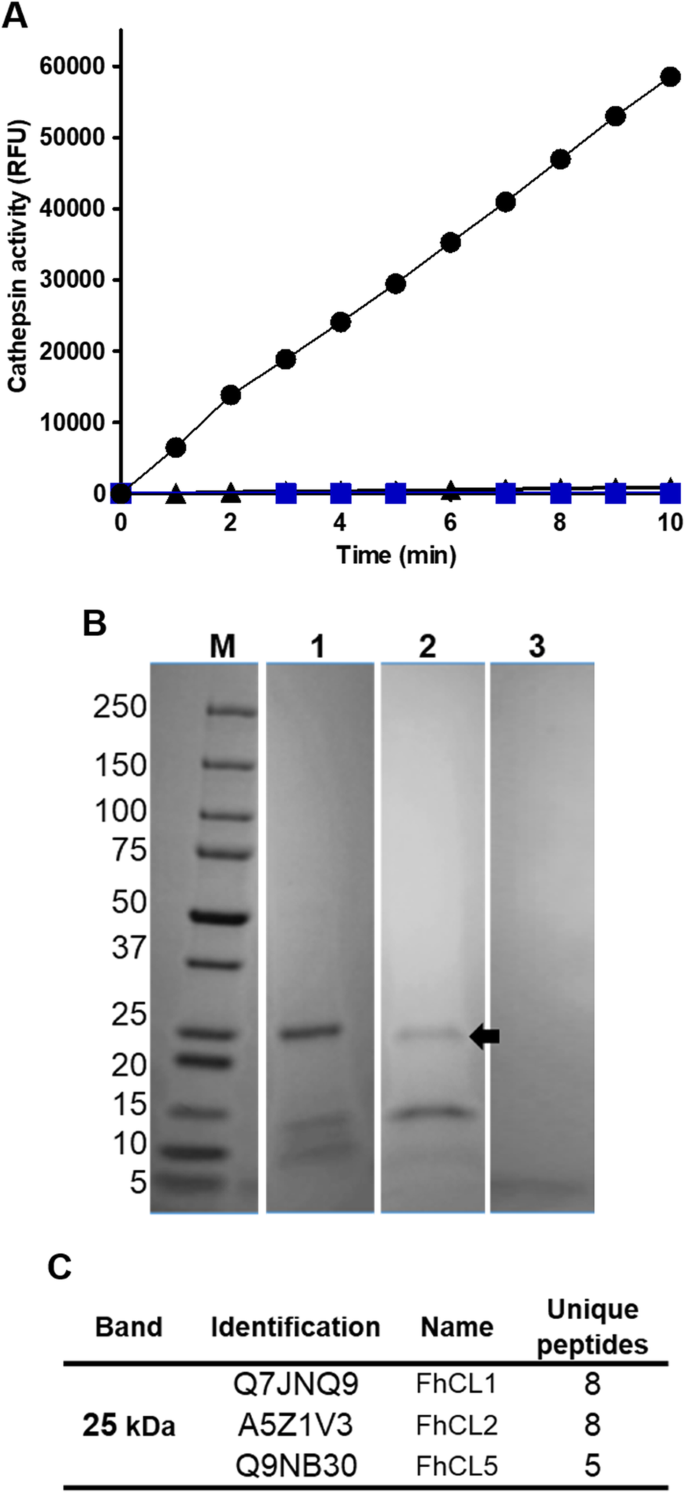
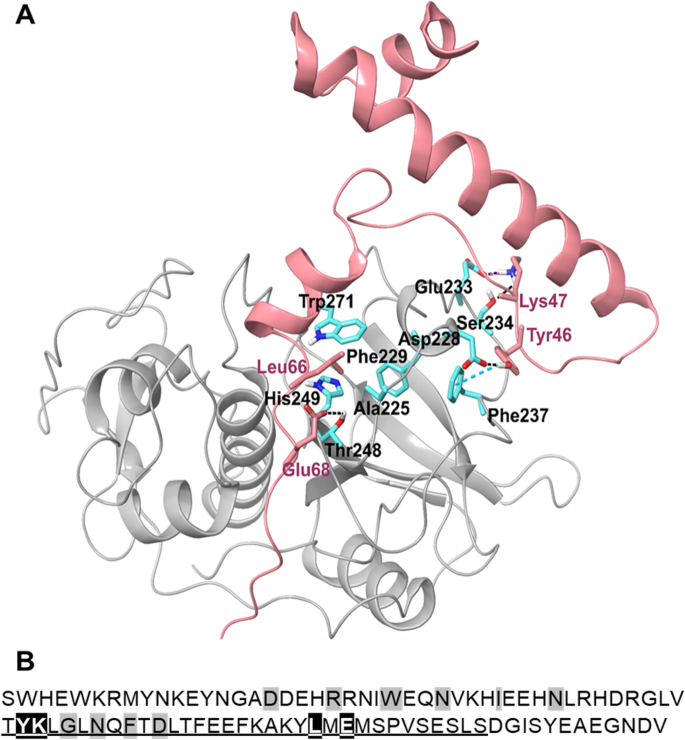
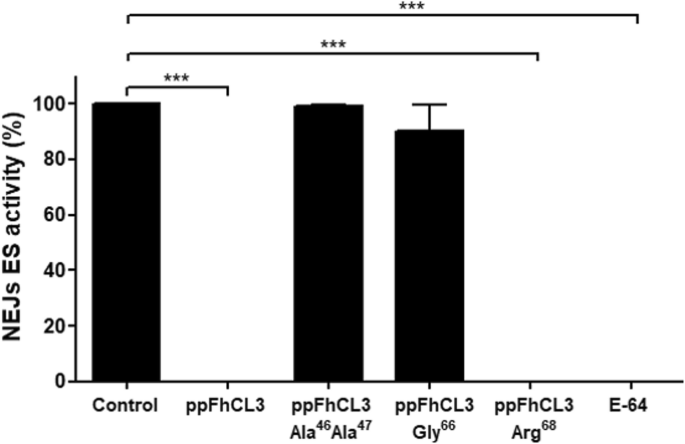
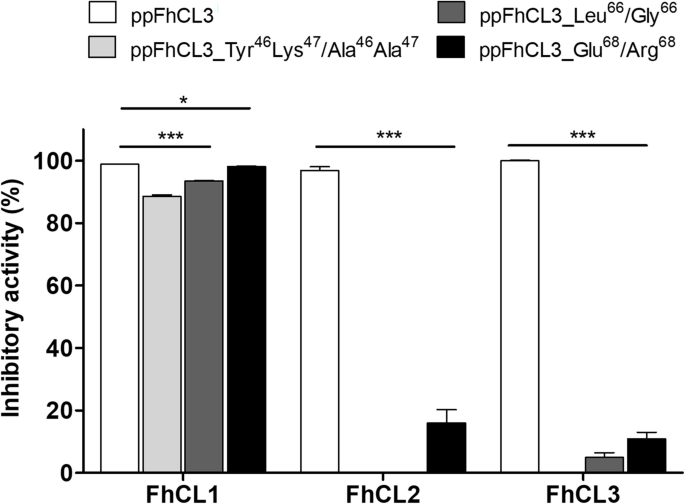
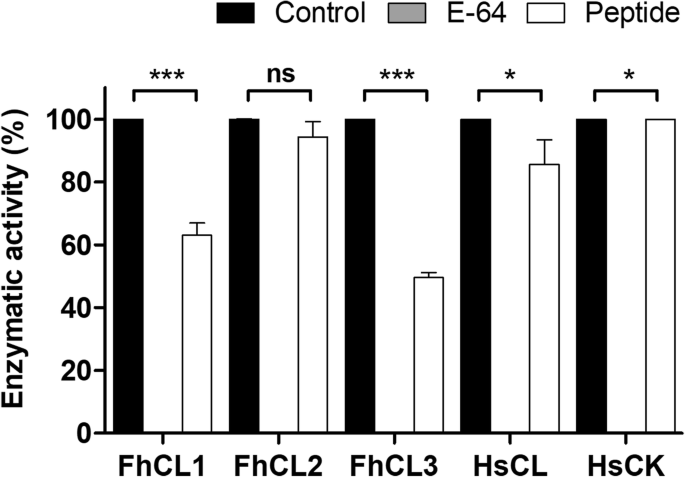
Immunolocalisation/immunoblotting studies show that the FhCL3 zymogen is produced and secreted by gastrodermal cells of the NEJs gut. A recombinant propeptide of FhCL3 (ppFhCL3) was shown to be a highly potent and selective inhibitor of native and recombinant F. hepatica FhCL3 peptidase, and other members of the cathepsin L family; inhibition constant (Ki) values obtained for FhCL1, FhCL2 and FhCL3 were 0.04 nM, 0.004 nM and < 0.002 nM, respectively. These values are at least 1000-fold lower than those Ki obtained for human cathepsin L (HsCL) and human cathepsin K (HsCK) demonstrating the selectivity of the ppFhCL3 for parasite cathepsins L. By exploiting 3-D structural data we identified key molecular interactions in the specific binding between the ppFhCL3 and FhCL3 mature domain. Using recombinant variants of ppFhCL3 we demonstrated the critical importance of a pair of propeptide residues (Tyr46Lys47) for the interaction with the propeptide binding loop (PBL) of the mature enzyme and other residues (Leu66 and Glu68) that allow the propeptide to block the active site. Conclusions: The FhCL3 peptidase involved in host invasion by F. hepatica is produced as a zymogen in the NEJs gut. Regulation of its activation involves specific binding sites within the propeptide that are interdependent and act as a "clamp-like" mechanism of inhibition. These interactions are disrupted by the low pH of the NEJs gut to initiate autocatalytic activation. Our enzyme kinetics data demonstrates high potency and selectivity of the ppFhCL3 for its cognate FhCL3 enzyme, information that could be utilised to design inhibitors of parasite cathepsin L peptidases.