Background
Histone methylation modifies the epigenetic state of target genes to regulate gene expression in the context of developmental and environmental changes. Previously, we used a positive genetic screen to identify an Arabidopsis mutant, cli186, which was impaired in carbon and light signaling. Here, we report a deletion of the Arabidopsis histone methyltransferase SDG8 in this mutant (renamed sdg8-5), which provides a unique opportunity to study the global function of a specific histone methyltransferase within a multicellular organism.
Conclusions
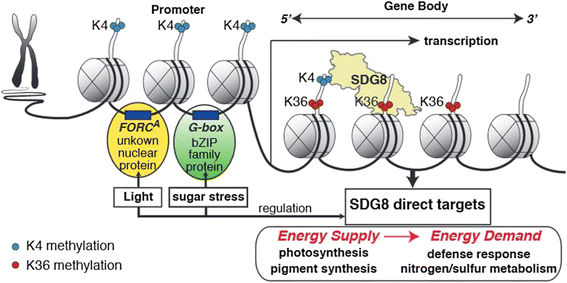
The histone methyltransferase SDG8 functions to regulate the H3K36 methylation of histones associated with gene bodies in Arabidopsis. The H3K36me3 mark in turn is associated with high-level expression of a specific set of light and/or carbon responsive genes involved in photosynthesis, metabolism and energy production.
Results
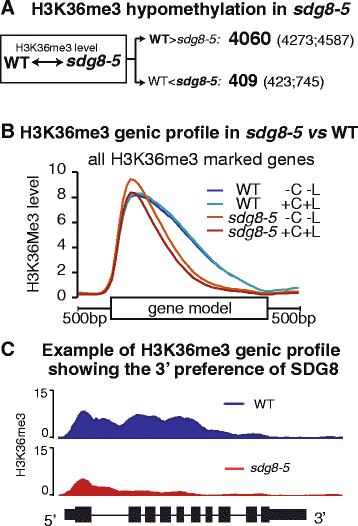
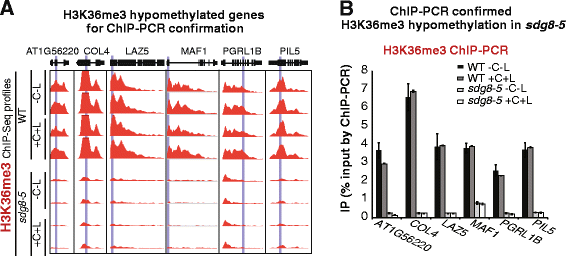
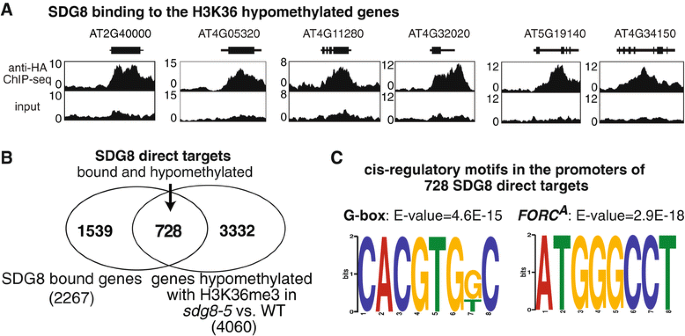
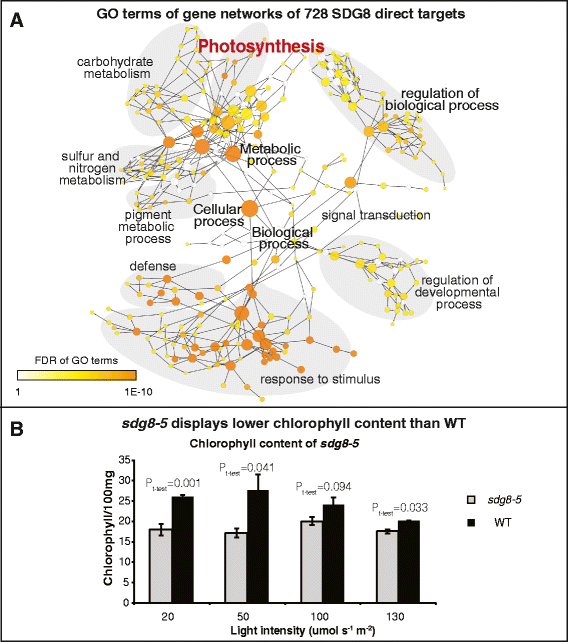
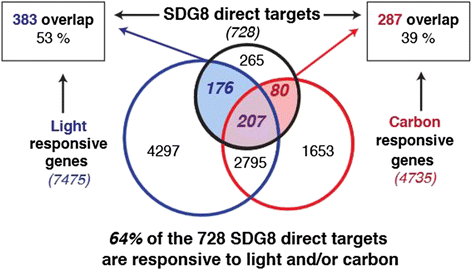
To assess the specific role of SDG8, we examine how the global histone methylation patterns and transcriptome were altered in the sdg8-5 deletion mutant compared to wild type, within the context of transient light and carbon treatments. Our results reveal that the sdg8 deletion is associated with a significant reduction of H3K36me3, preferentially towards the 3' end of the gene body, accompanied by a reduction in gene expression. We uncover 728 direct targets of SDG8 that have altered methylation in the sdg8-5 mutant and are also bound by SDG8. As a group, this set of SDG8 targets is enriched in specific biological processes including defense, photosynthesis, nutrient metabolism and energy metabolism. Importantly, 64% of these SDG8 targets are responsive to light and/or carbon signals. Conclusions: The histone methyltransferase SDG8 functions to regulate the H3K36 methylation of histones associated with gene bodies in Arabidopsis. The H3K36me3 mark in turn is associated with high-level expression of a specific set of light and/or carbon responsive genes involved in photosynthesis, metabolism and energy production.