1. 领域背景与文献引入
文献英文标题:Telomere-to-telomere assembly of a fish Y chromosome reveals the origin of a young sex chromosome pair;发表期刊:Genome Biology;影响因子:13.583(2021年);研究领域:鱼类性别染色体进化、单倍型基因组组装、着丝粒周区异染色质功能。
性别染色体进化是生命科学领域的核心研究方向之一,其起源于普通常染色体对,重组抑制的建立是驱动性别染色体分化的关键步骤。理论研究表明,当性拮抗多态性靠近性别决定位点时,自然选择会倾向于维持该区域的重组抑制,进而推动性别连锁区的形成与扩张。在哺乳动物、鸟类等类群中,性别染色体已高度分化,Y/W染色体呈现基因贫乏、重复序列富集的特征;而在鱼类、两栖类等脊椎动物中,性别染色体常为同态,起源时间较近,是研究性别染色体早期分化机制的理想模型。然而,由于年轻性别染色体的X/Y(或Z/W)序列差异极小且富含重复序列,传统短读长测序和错误率较高的长读长技术难以准确组装单倍型基因组,导致其早期进化机制的研究存在诸多空白。现有研究多聚焦于通过结构变异(如倒位、易位)建立重组抑制的性别染色体,对预存低重组区(如着丝粒周区)起源的性别连锁区研究较少,尤其是在动物类群中缺乏直接实验证据。
本文针对这一研究空白,采用高准确率的PacBio HiFi长读长测序结合Hi-C染色质构象捕获技术,完成了棘鳅(Mastacembelus armatus)的单倍型分辨率染色体级基因组组装,首次在鱼类中端粒到端粒解析了Y染色体的完整结构,揭示了年轻性别连锁区起源于着丝粒周区的预存低重组异染色质,无需依赖结构变异,为动物性别染色体的早期进化机制提供了新的实验证据。
2. 文献综述解析
本文作者从性别染色体进化的核心机制、基因组组装技术的迭代、年轻性别染色体研究的挑战三个维度对现有研究进行了系统评述,明确了领域内的研究热点与未解决问题,为自身研究的创新定位奠定了基础。
现有研究的关键结论显示,性别染色体的进化遵循从常染色体对到分化完全性染色体的路径,重组抑制是驱动这一过程的核心力量,性拮抗选择、遗传搭车、背景选择等机制共同推动了Y/W染色体的退化。在技术方法层面,早期短读长测序技术无法组装复杂的重复序列区域,普通长读长(如PacBio CLR、Nanopore)错误率较高,难以区分近缘的X/Y单倍型;trio-binning方法需要家系样本,限制了非模式生物的应用;而HiFi长读长测序技术准确率超过99.5%,结合Hi-C数据可实现无需家系的单倍型基因组组装,为年轻性别染色体的研究提供了关键技术支撑。然而,现有研究仍存在局限性:多数年轻性别染色体的研究集中在存在结构变异的类群,对预存低重组区起源的性别连锁区研究主要集中在植物中,动物类群中的直接证据不足;同时,鱼类作为性别染色体多样性最高的类群之一,其年轻性别染色体的起源机制尚未被充分解析。
本文的创新价值在于,首次在鱼类中通过端粒到端粒的Y染色体组装,直接证明了年轻性别连锁区可起源于着丝粒周区的预存低重组异染色质,无需依赖结构变异,补充了动物性别染色体早期进化的机制模型;同时,利用单倍型基因组解析了着丝粒周区异染色质的结构与功能特征,为低重组区在性别染色体起源中的作用提供了新的视角。
3. 研究思路总结与详细解析
本文的研究目标是构建棘鳅的单倍型分辨率染色体级基因组,解析年轻性别染色体的起源与重组抑制机制;核心科学问题为年轻性别连锁区的起源是否依赖结构变异,着丝粒周区的预存低重组环境如何促进性别连锁区的形成;技术路线遵循“基因组组装→核型与功能元件鉴定→性别连锁区定位→候选基因筛选→机制验证”的闭环逻辑,通过多组学技术结合的方法系统解析了年轻性别染色体的进化过程。
3.1 单倍型分辨率染色体级基因组组装
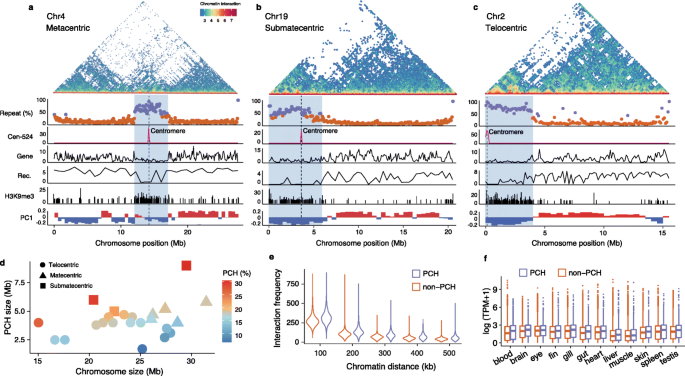
实验目的是获得高质量的棘鳅单倍型基因组,准确区分X和Y染色体,为后续性别染色体分析提供基础。方法细节:提取雄性棘鳅肌肉组织DNA,采用PacBio HiFi测序技术生成约30G的测序数据,k-mer分析显示HiFi reads的错误率仅为0.086%,并估计基因组大小为600.1Mb;先构建单倍型共识序列,再结合HiFi reads自身的分型信息与Hi-C染色质相互作用数据,利用Whatshap和hapCUT2软件进行单倍型分型,将reads分配到hap-X和hap-Y两个单倍型,分别组装得到两个包含24条染色体的单倍型基因组;通过将已报道的雄性特异性标记映射到单倍型基因组,确定Y染色体所属的单倍型。结果解读:hap-Y基因组的质量更优,所有24条染色体的缺口数均小于10,平均缺口数为5.5,其中Y染色体实现了端粒到端粒的无缺口组装;hap-X基因组的平均缺口数为6.4;基因组共线性分析显示X和Y染色体之间无大规模结构变异,仅在端粒区域存在少量倒位;系统发育分析表明棘鳅与亚洲沼泽鳗的分化时间约为3600万年前。
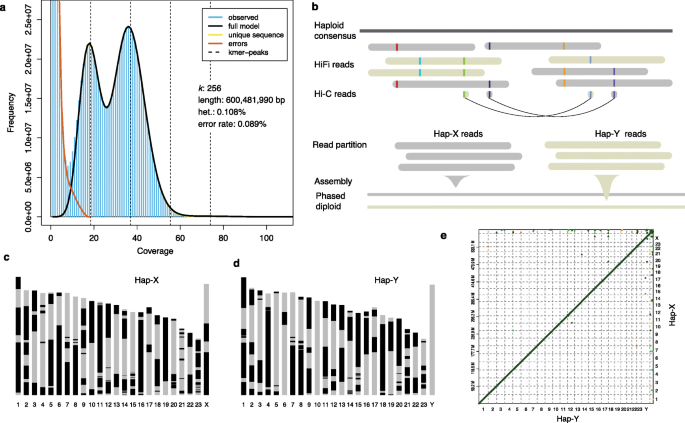
产品关联:文献未提及具体实验产品,领域常规使用PacBio Sequel II测序平台、Hi-C建库试剂盒、基因组组装软件(peregrine、3D-DNA)。
3.2 核型进化与着丝粒卫星序列鉴定
实验目的是验证基因组组装的染色体级准确性,鉴定棘鳅的着丝粒和端粒特异性卫星序列,为着丝粒周区异染色质分析提供基础。方法细节:通过染色体核型分析确定棘鳅的二倍体染色体数为48,根据染色体形态分为5个中着丝粒、3个亚中着丝粒和16个端着丝粒染色体;利用RepeatModeler和RepeatMasker软件进行重复序列注释,筛选出丰度最高的两种卫星序列(Cen-524,524bp;Tel-190,190bp);通过荧光原位杂交技术,将Cen-524和Tel-190的探针分别标记为红色(Cy5-dUTP)和绿色(FITC-dUTP),与中期染色体杂交验证其定位。结果解读:核型分析结果与基因组组装的染色体数目一致,染色体形态分类与基因组的着丝粒位置预测相符;Cen-524卫星序列仅在每条染色体的着丝粒区域出现一个或零个峰,端着丝粒染色体中位于染色体末端,中着丝粒和亚中着丝粒染色体中位于中部,是着丝粒的特异性标记;Tel-190卫星序列仅位于染色体末端,中着丝粒染色体中有时位于两端,与端粒相关;荧光原位杂交结果与基因组序列的定位完全一致,验证了基因组组装的准确性;着丝粒的平均组装长度为50.5kb,但大部分着丝粒的组装不完整,端粒的保守基序(TTAGGG)n未被组装,但荧光原位杂交显示其与Tel-190共存。
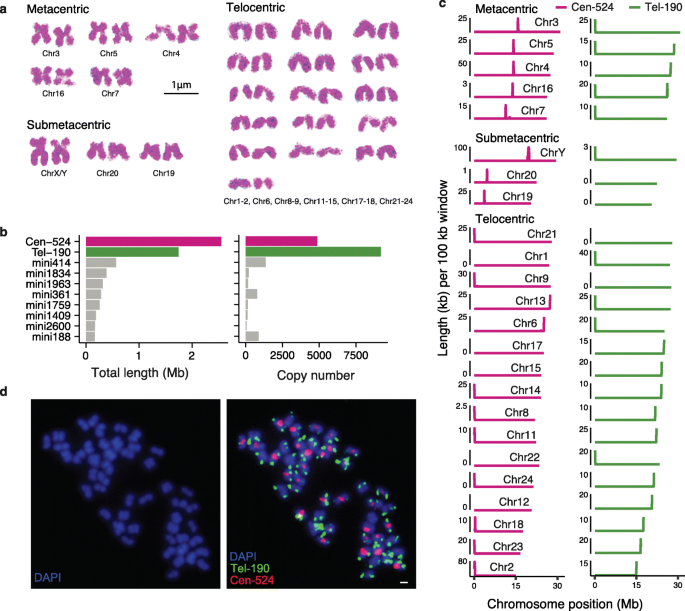
产品关联:实验所用关键产品:Roche Nick Translation Mix探针标记试剂盒、Olympus BX63荧光显微镜、DAPI染色液(Vector Laboratories)。
3.3 年轻性别染色体与性别连锁区定位
实验目的是定位棘鳅的性别连锁区(SLR),分析其序列特征与重组抑制机制,探讨其起源模式。方法细节:对10尾雄性和10尾雌性棘鳅进行全基因组重测序,利用GATK软件进行SNP calling与过滤,通过EMMAX软件进行全基因组关联分析(GWAS)筛选与性别相关的遗传变异;计算X和Y染色体在100kb窗口内的序列相似性,统计每50kb窗口内的雄性特异性SNP密度;结合Hi-C染色质区室分析、CUT&Tag技术检测的H3K9me3异染色质修饰、重复序列含量分析,解析SLR的染色质特征。结果解读:GWAS分析显示,约7Mb的Y染色体区域与性别显著关联(log10转换后的p值显著富集),该区域跨越着丝粒,几乎完全嵌入着丝粒周异染色质;根据雄性特异性SNP密度和X/Y序列相似性,SLR可分为两个区域:R1区域(3.0Mb)包含67个蛋白编码基因,雄性特异性SNP密度高,X/Y序列相似性显著低于R2区域(Wilcoxon sum rank test,P=0.013);R2区域(3.9Mb)基因贫乏,仅含22个基因;X和Y染色体的SLR序列与基因含量几乎相同,但均富含重复序列、具有H3K9me3修饰、属于沉默的B染色质区室,显示出零或低重组的特征;染色质相互作用分析显示,着丝粒周区的染色质远程相互作用更频繁,表明其折叠更紧密。
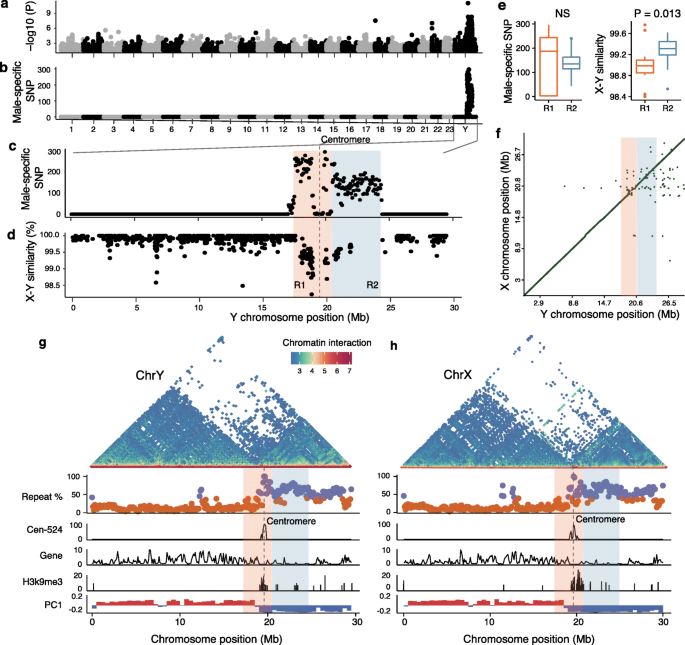
产品关联:文献未提及具体实验产品,领域常规使用Illumina HiSeq Xten重测序平台、CUT&Tag试剂盒、GWAS分析软件(EMMAX)。
3.4 候选性别决定基因筛选与表达模式分析
实验目的是从SLR中筛选候选性别决定基因,分析其表达模式与功能特征,验证其在性别决定中的作用。方法细节:利用HiSat2和featureCounts软件对5尾雄性、5尾雌性、5尾间性棘鳅的性腺RNA-seq数据进行比对与定量,以TPM(转录本每百万)值衡量基因表达水平;筛选SLR中具有性别特异性表达的基因,重点分析其结构域与功能;通过系统发育分析确定候选基因的进化地位。结果解读:SLR中的基因未表现出普遍的组织特异性表达,但HMGN6和SYCE3两个基因的表达模式具有显著性别偏向性;HMGN6在精巢和早期间性腺中的表达水平相似,而在卵巢中的表达水平极低,其编码的蛋白含有HMG结构域,与哺乳动物的性别决定基因SRY的结构域同源,是潜在的性别决定基因;SYCE3仅在精巢中高表达,在间性腺的晚期阶段也有表达,其已知功能是参与减数分裂的联会复合体形成,可能是雄性有益基因;系统发育分析显示HMGN6仅存在于硬骨鱼类中,与四足动物的HMGN5同源。
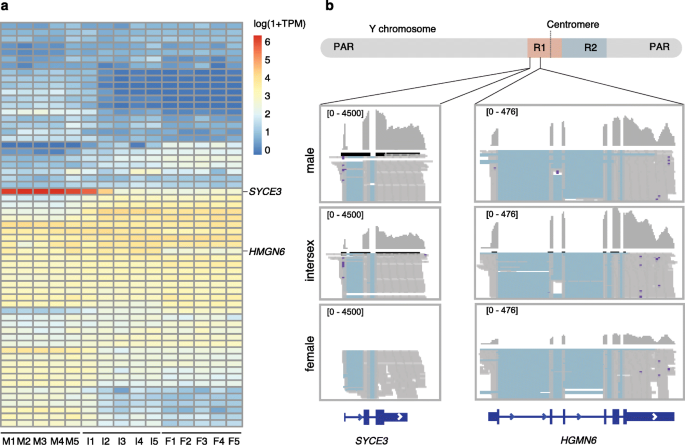
产品关联:实验所用关键产品:NEBNext Ultra RNA Library Prep Kit建库试剂盒、Illumina HiSeq Xten测序平台、转录组分析软件(HiSat2、featureCounts)。
3.5 着丝粒周异染色质区的结构与功能特征分析
实验目的是分析常染色体着丝粒周异染色质(PCH)的特征,探讨其与SLR起源的相关性,验证预存低重组区的作用。方法细节:通过重复序列含量、基因密度、H3K9me3修饰、染色质区室、重组率等指标,在常染色体上界定着丝粒周异染色质的范围;比较PCH与非PCH区域的染色质相互作用频率、基因表达水平与广度;利用ReLERNN软件估计常染色体的重组率。结果解读:常染色体的PCH大小约为4.2Mb,与染色体大小呈弱的非显著正相关(Pearson’s r=0.34,P=0.11),较小的染色体(如端着丝粒、亚中着丝粒染色体)的PCH占比更高;PCH区域的重复序列含量超过50%,基因密度低,具有H3K9me3修饰,属于沉默的B染色质区室,重组率显著低于非PCH区域;PCH内的染色质远程相互作用频率显著高于非PCH区域(P<2.2e-16,Wilcoxon rank sum test),表明其染色质折叠更紧密;意外的是,PCH区域的基因表达水平与广度显著高于非PCH区域(P=2.279e-11,Wilcoxon rank sum test),提示H3K9me3修饰对PCH内基因的抑制作用有限,可能存在其他表观修饰调控基因表达。
产品关联:文献未提及具体实验产品,领域常规使用Hi-C分析软件(Juicer、Juicebox)、重组率估计软件(ReLERNN)。
4. Biomarker研究及发现成果解析
本文涉及的Biomarker包括棘鳅的性别连锁区(SLR)分子标记与候选性别决定基因HMGN6,通过多组学技术结合的方法完成了筛选与验证,为鱼类性别鉴定与性别决定机制研究提供了新的分子靶点。
Biomarker定位与筛选逻辑
性别连锁区是位于Y染色体着丝粒周区的~7Mb区域,其筛选与验证逻辑为:通过10雄10雌的全基因组重测序进行GWAS分析,定位到与性别显著关联的区域;结合雄性特异性SNP密度、X/Y序列相似性分析,确定该区域为完全性别连锁区;通过染色质特征分析验证其低重组的异染色质属性。候选性别决定基因HMGN6的定位逻辑为:在SLR内筛选具有性别偏向性表达的基因;结合结构域分析(HMG结构域)与表达模式(精巢/早期间性腺特异性表达),确定其为候选性别决定基因;通过系统发育分析验证其进化保守性。
研究过程与验证数据
性别连锁区来源于棘鳅Y染色体的着丝粒周异染色质,验证方法包括GWAS分析(log10转换后的p值在该区域超过10,显示极强的性别关联信号)、雄性特异性SNP密度分析(每50kb窗口的SNP数在SLR内显著高于其他区域)、X/Y序列相似性分析(100kb窗口的序列相似性在R1区仅为~99%,显著低于R2区);HMGN6的验证方法为性腺RNA-seq定量,其在精巢中的TPM值显著高于卵巢(n=5,P<0.01),在早期间性腺中的TPM值与精巢相近;性别连锁区的特异性表现为仅在雄性个体中存在雄性特异性变异,雌性个体中无对应变异;敏感性方面,GWAS分析的p值在SLR区域的富集程度远高于基因组其他区域,可作为性别鉴定的可靠分子标记。
核心成果与创新价值
本文鉴定的性别连锁区是首次在鱼类中发现的起源于着丝粒周预存低重组区的年轻性别连锁区,其创新性在于证明了动物性别连锁区的起源无需依赖结构变异,预存低重组异染色质即可为性别连锁区的形成提供基础,补充了性别染色体早期进化的机制模型;HMGN6作为候选性别决定基因,首次在硬骨鱼类中发现HMG家族基因参与性别决定,为鱼类性别决定的分子机制研究提供了新的方向;统计学结果显示,X/Y序列相似性在R1区与R2区的差异显著(Wilcoxon sum rank test,P=0.013),PCH与非PCH区域的基因表达广度差异显著(P=2.279e-11),为结论提供了坚实的统计支撑。推测:HMGN6可能通过结合染色质调控性别决定相关基因的表达,其具体功能需通过基因敲除、过表达等功能实验进一步验证。