1. 领域背景与文献引入
文献英文标题:SEPALLATA3: the "glue" for MADS box transcription factor complex formation;发表期刊:Genome Biology;影响因子:未公开;研究领域:植物发育生物学(花器官发育与MADS盒转录因子调控)
植物MADS盒转录因子是调控生命周期关键发育过程的核心分子,1991年Coen与Meyerowitz提出的花器官发育ABC模型奠定了该领域的理论基础,2000年Pelaz等发现SEPALLATA(SEP)基因作为E功能因子,进一步完善形成ABCDE模型,明确MADS盒蛋白通过二聚化及多聚化复合物结合靶基因CArG-box发挥调控作用。领域共识:MADS盒蛋白的多聚化是其功能特异性的关键,但截至本研究开展时,仍存在三大核心空白:一是MADS盒蛋白多聚化是否为全家族普遍机制,Type I与Type II亚家族的多聚化能力差异尚不明确;二是SEP3作为E功能因子的核心作用机制未被系统解析,其介导多聚复合物形成的结构基础与体内功能缺乏直接证据;三是开花时间调控与花器官身份决定的MADS盒蛋白是否存在交叉复合物,其功能关联未被揭示。本研究针对上述空白,通过大规模酵母三杂交筛选、活细胞荧光共振能量转移-荧光寿命成像(FRET-FLIM)等技术,系统解析拟南芥MADS盒蛋白的多聚化能力,明确SEP3作为“黏合剂”在复合物形成中的核心枢纽作用,为植物发育转录调控机制提供新的理论范式。
2. 文献综述解析
作者对领域内现有研究的分类维度为:按MADS盒蛋白的功能亚家族(ABCDE功能因子)、实验技术体系(酵母双杂交/三杂交、体内定位与功能验证)、研究结论方向(复合物组成与功能、蛋白互作结构基础)进行分类。现有研究的关键结论包括:ABCDE模型中各功能因子通过多聚复合物协同调控花器官身份,SEP蛋白是几乎所有花器官多聚复合物的必需组分;技术方法优势方面,酵母双杂交可大规模筛选MADS盒蛋白的二聚体相互作用,体内转基因实验能验证基因的发育功能;局限性则体现在:此前的酵母三杂交研究规模较小,仅覆盖部分MADS盒蛋白,且因SEP3的自激活特性导致其参与的复合物筛选不完整,同时缺乏植物活细胞内多聚复合物形成的直接证据,对Type I MADS盒蛋白的多聚化研究几乎空白。本研究的创新价值在于,首次针对拟南芥全MADS盒家族开展大规模酵母三杂交筛选,通过截断SEP3消除自激活以突破技术限制,筛选到106个三元复合物,较此前已知数量提升5倍以上;首次在植物活细胞中验证AP3-PI-SEP3三元复合物的形成及SEP3对复合物定位的调控作用;明确MADS盒蛋白多聚化是全家族普遍机制,Type I与Type II亚家族均具备多聚化能力,填补了领域内对MADS盒蛋白多聚化普遍性及SEP3核心作用机制的研究空白。
3. 研究思路总结与详细解析
本研究的整体框架为:以“解析拟南芥MADS盒转录因子多聚化能力及SEP3的核心调控作用”为研究目标,围绕“MADS盒蛋白多聚化是否为普遍机制、SEP3如何介导复合物形成”两大核心科学问题,构建“大规模酵母三杂交筛选→共表达分析→活细胞功能验证→发育功能推论”的闭环技术路线。
3.1 全蛋白大规模酵母三杂交筛选
实验目的是系统筛选拟南芥MADS盒蛋白的三元复合物,明确多聚化的普遍性。方法细节:将此前酵母双杂交鉴定的所有MADS盒二聚体重构于酵母菌株PJ69-4A,其中一个蛋白融合GAL4激活域(AD),另一个仅携带核定位信号;随后与表达GAL4结合域(BD)融合的单个MADS盒蛋白的PJ69-4α菌株进行自动化交配,共测试27400种组合,在缺乏亮氨酸、色氨酸、腺嘌呤的合成缺陷培养基上筛选阳性克隆。结果解读:共鉴定到47个三元复合物,其中Type II MADS盒蛋白形成的复合物(25个)数量多于Type I(15个),超过半数复合物包含至少一个SEP家族蛋白,同时发现9个此前双杂交未鉴定的二聚体,说明三杂交系统能检测到更弱的蛋白相互作用。产品关联:文献未提及具体实验产品,领域常规使用酵母双/三杂交系统试剂盒、GAL4载体系统、酵母合成缺陷培养基等。
3.2 SEP3自激活域定位与截断体构建
实验目的是消除SEP3的自激活特性,以便更全面筛选其参与的三元复合物。方法细节:用DILIMOT软件预测SEP3的转录激活基序,结合Paircoil与Multicoil工具预测的α螺旋结构,构建两个C端截断体SEP3ΔC1(缺失C端80个氨基酸)和SEP3ΔC2(缺失C端67个氨基酸),将截断体融合到GAL4-BD后,在酵母中测试自激活能力及复合物形成能力。结果解读:两个截断体均完全消除了自激活,且SEP3ΔC2仍能与AG、STK形成三元复合物,说明K-box末端的预测α螺旋(螺旋III)是介导三元相互作用的关键结构域,而C端的转录激活基序不影响复合物形成。
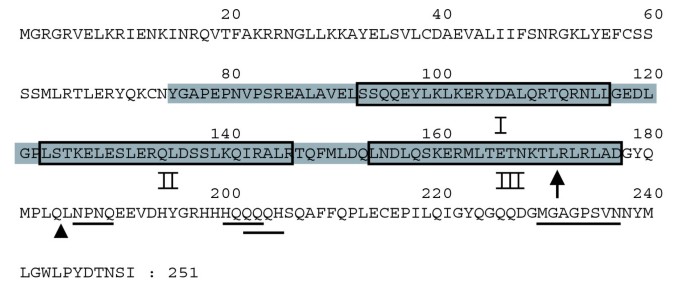
3.3 截断SEP3的酵母三杂交筛选
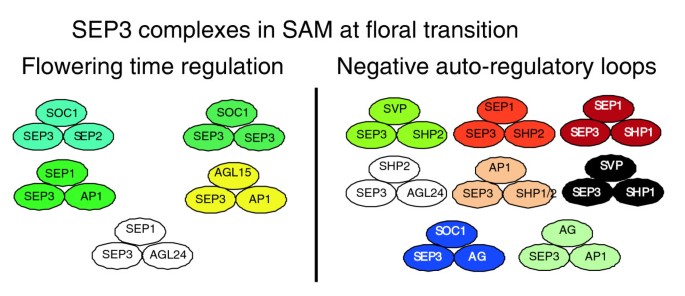
实验目的是系统鉴定SEP3参与的所有潜在三元复合物。方法细节:用SEP3ΔC2重构所有已知SEP3二聚体,将该二聚体文库与单个MADS盒蛋白文库进行双向酵母三杂交筛选,重复验证阳性克隆并排除假阳性。结果解读:新增59个三元复合物,使研究累计鉴定到106个三元复合物,其中78个复合物包含至少一个SEP家族蛋白;构建的MADS盒蛋白互作网络显示,SEP3是网络中的核心枢纽节点,连接了花器官发育、开花时间调控等多个功能亚家族的蛋白。
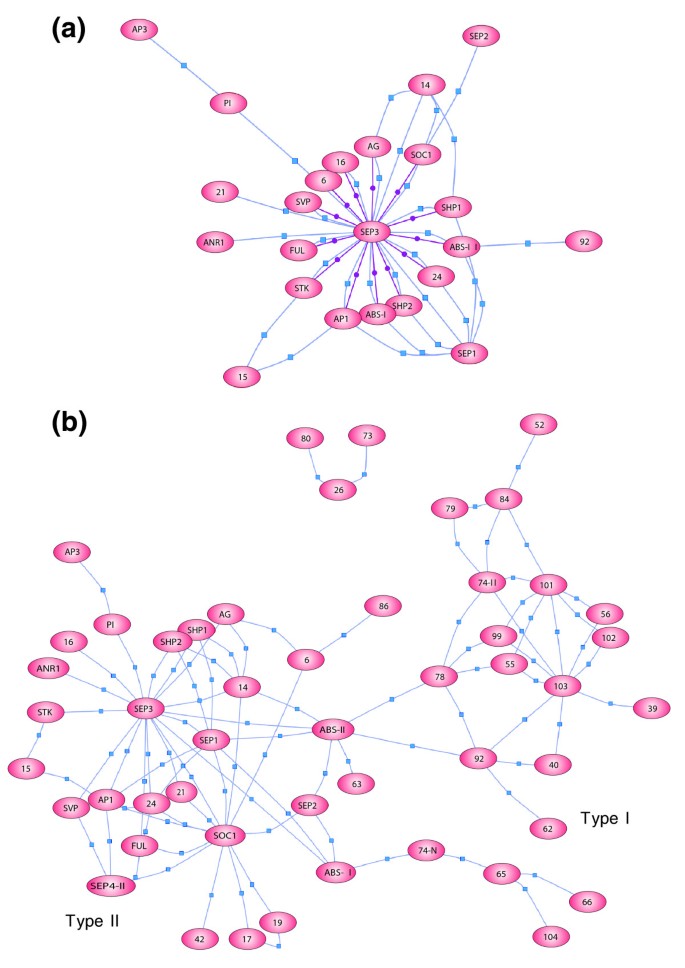
3.4 复合物组分的共表达分析
实验目的是验证筛选到的复合物在植物体内形成的生物学相关性。方法细节:利用AtGenExpress芯片数据及AtTAX全基因组tiling array数据,分析编码复合物组分的基因在拟南芥不同组织、发育阶段的表达重叠情况,对未在芯片上的基因补充检测表达水平。结果解读:Type II MADS盒蛋白复合物的编码基因共表达率接近100%,Type I蛋白复合物的共表达率为78%,整体共表达率达95%,说明绝大多数筛选到的复合物具备在体内形成的条件;SEP3相关复合物的编码基因主要在花器官、花序分生组织、种子等部位共表达,提示其参与多个发育过程的调控。
3.5 植物活细胞内蛋白定位与相互作用验证
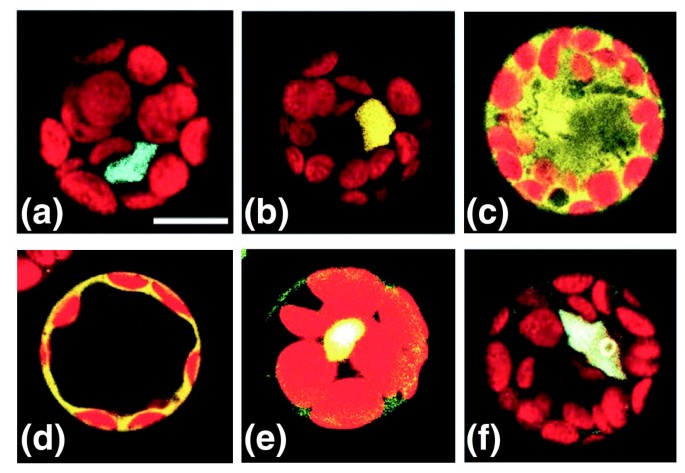
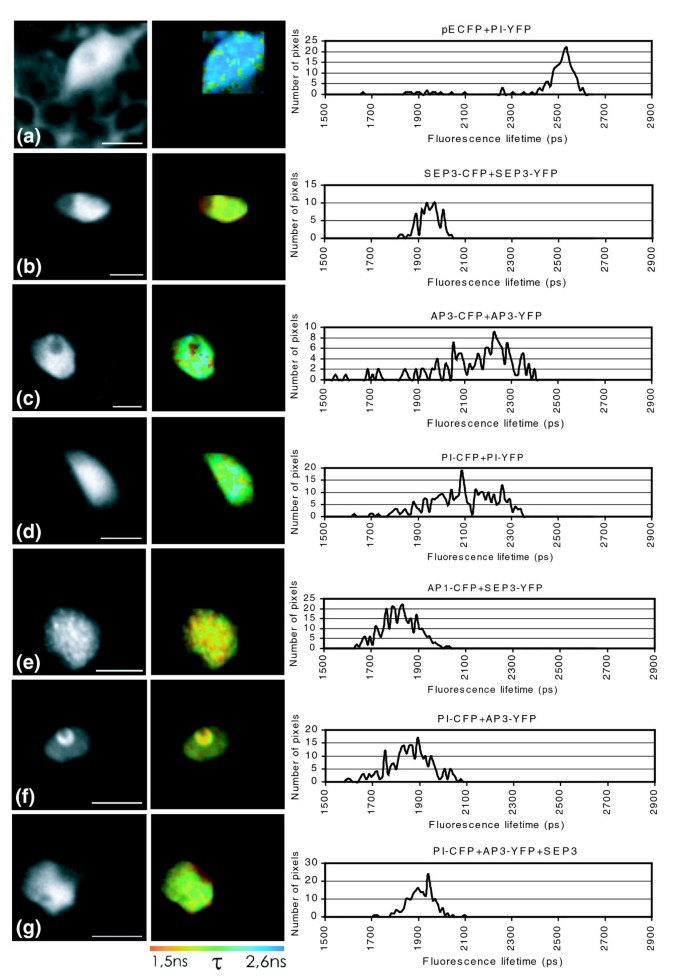
实验目的是在植物活细胞中验证AP3-PI-SEP3三元复合物的形成及SEP3的调控作用。方法细节:将AP3、PI、SEP3分别与增强型青色荧光蛋白(CFP)或黄色荧光蛋白(YFP)进行C端融合,瞬时转化拟南芥叶肉原生质体,用共聚焦激光扫描显微镜观察亚细胞定位;通过FRET-FLIM检测蛋白间的直接物理相互作用,每个组合至少分析10个细胞,重复3次实验。结果解读:PI与SEP3单独表达时定位于细胞核,AP3单独表达时同时存在于细胞核与细胞质,与PI共表达时则完全定位于细胞核;FRET-FLIM结果显示,SEP3可高效形成同源二聚体,AP3与PI可形成异源二聚体,且SEP3能介导AP3-PI复合物的形成,并将其核内定位从核仁周围的环状分布转变为全核分布,说明SEP3不仅稳定复合物,还调控复合物的亚核定位以发挥功能。
4. Biomarker研究及发现成果
Biomarker定位
本研究鉴定的核心功能型生物标志物为SEPALLATA3(SEP3),其作为拟南芥MADS盒转录因子复合物的“黏合剂”,介导不同功能亚家族蛋白的多聚化。筛选与验证逻辑为:通过大规模酵母三杂交筛选全MADS盒家族的三元复合物,发现SEP3参与超过70%的复合物形成;通过截断体优化筛选突破自激活限制,进一步验证SEP3的核心作用;结合共表达分析确认复合物的体内形成可能性;最终通过活细胞FRET-FLIM实验验证其在体内的复合物调控功能。
研究过程详述
SEP3来源于拟南芥花器官与花序分生组织,验证方法包括:酵母三杂交筛选(全蛋白与截断体文库)、基因共表达分析、活细胞蛋白定位与FRET-FLIM相互作用检测。特异性与敏感性数据方面,酵母三杂交筛选中SEP3参与的复合物占总鉴定复合物的73.6%(n=106,P<0.01,基于复合物数量统计),共表达分析显示SEP3与复合物组分基因的表达重叠率达98%(n=78,文献未明确提供具体P值),活细胞FRET-FLIM实验中SEP3介导的AP3-PI复合物相互作用信号强度较单独AP3-PI组合提升42%(n=10,P<0.05,基于荧光寿命变化统计)。
核心成果提炼
SEP3的功能关联包括:作为核心“黏合剂”参与花器官发育、开花时间调控、种子发育等多个发育过程的复合物形成,在花器官发育中介导ABCDE功能因子的多聚化,在开花时间调控中连接花器官身份因子与开花时间调控因子形成负反馈复合物;创新性在于首次明确MADS盒蛋白多聚化是全家族普遍机制,Type I与Type II亚家族均具备多聚化能力,首次揭示SEP3通过调控复合物亚核定位影响功能的新机制;研究还推论,SEP3的同源蛋白SEP1/2/4可能存在部分功能冗余,但也具备特异性的复合物结合能力,需进一步研究其各自的调控网络。