1. 领域背景与文献引入
文献英文标题:Mutational analysis of βCOP (Sec26p) identifies an appendage domain critical for function;发表期刊:BMC Cell Biology;影响因子:未公开;研究领域:细胞生物学-囊泡运输与COPI复合物功能。
真核细胞的囊泡运输系统负责在分泌和内吞途径的膜性细胞器之间转运货物,是维持细胞稳态的核心机制之一。COPI包被囊泡(COPI-coated vesicle)作为逆向运输的关键载体,主要介导从顺式高尔基体到内质网的货物回流,同时参与高尔基体内部的物质转运,其功能异常会导致蛋白定位紊乱、分泌缺陷等多种细胞病理表型。领域共识:COPI复合物由7个保守的coatomer亚基组成,可分为F-COPI(含β、γ、δ、ζCOP)和B-COPI(含α、β"、εCOP)两个亚复合物,其中γCOP的附加结构域(appendage domain)已被证实与adaptin家族蛋白的结构同源,通过FxxxW基序参与配体结合与信号调控,但βCOP(酿酒酵母中同源物为Sec26p)是否存在具有功能的同源附加结构域,以及该结构域在COPI囊泡运输中的具体作用机制,尚未得到明确验证,这一空白限制了对COPI复合物调控网络的完整理解。
针对这一研究空白,本研究通过生物信息学预测、遗传突变筛选及功能验证相结合的方法,系统探究了βCOP附加结构域的存在性与功能必需性,并进一步揭示了其与ArfGAP蛋白Glo3p的非催化依赖型相互作用,为COPI囊泡运输的调控机制提供了新的关键证据。
2. 文献综述解析
作者从COPI复合物的结构组成、adaptin家族附加结构域的功能特征、ArfGAP蛋白的调控作用三个维度,系统梳理了领域内的研究进展,明确了现有研究的核心结论与未解决问题,为本文的研究设计奠定了逻辑基础。
现有研究已证实COPI复合物的亚基组成与基本功能,γCOP附加结构域与AP2复合物的附加结构域具有高度结构同源性,尽管一级序列一致性仅为12%-16%,但二者均包含保守的FxxxW基序,该基序在AP2中负责招募调控网格蛋白包被囊泡组装的配体;ArfGAP蛋白作为小GTP酶Arf1的负调控因子,不仅参与COPI囊泡的脱包被过程,还在货物分选与膜曲率感知中发挥作用,其中Glo3p与Gcs1p在逆向运输中具有功能冗余性,共同构成必需的调控对。现有研究的技术优势在于通过结构生物学与遗传学手段明确了γCOP的功能,但局限性在于未对βCOP的潜在附加结构域进行系统验证,且对ArfGAP与βCOP的相互作用机制缺乏深入解析,部分研究仅停留在遗传相互作用层面,未明确具体的结构与功能关联。
与现有研究相比,本研究的核心创新点在于首次通过结构预测与功能缺失实验,明确βCOP的羧基端存在adaptin家族样附加结构域,且该结构域的平台亚域是酵母存活与COPI功能必需的;同时首次揭示Glo3p通过其羧基端的ISS基序而非催化结构域与βCOP相互作用,证明ArfGAP蛋白具有独立于GTP酶激活活性的调控功能,这一发现填补了COPI复合物调控网络中βCOP功能机制的空白,为囊泡运输领域的研究提供了新的方向。
3. 研究思路总结与详细解析
本研究采用“结构预测→功能验证→突变体筛选→调控因子鉴定”的闭环研究逻辑,以明确βCOP附加结构域的存在性与功能机制为核心科学问题,通过生物信息学、分子遗传学、细胞生物学等多维度技术手段,系统验证了βCOP附加结构域的功能必需性,并揭示了其与Glo3p的非催化依赖型相互作用。
3.1 βCOP(Sec26p)附加结构域的生物信息学预测
实验目的是通过结构生物学预测工具,验证Sec26p的羧基端是否具有与adaptin家族同源的附加结构域折叠特征。方法细节为使用PSIPRED二级结构预测服务器分析Sec26p的全序列二级结构,同时利用mGenTHREADER折叠识别工具对Sec26p的羧基端序列进行同源折叠匹配,并通过ClustalX软件将Sec26p与αAP2、γCOP的附加结构域进行多序列比对。结果解读:PSIPRED预测显示Sec26p的730-840位氨基酸为连续β折叠结构,对应adaptin家族附加结构域的免疫球蛋白样β-三明治亚域,840-973位为α螺旋与β折叠混合结构,对应平台亚域;mGenTHREADER分析显示Sec26p的700-973位序列与牛αAP2附加结构域的折叠匹配度为“Certain”(p<0.0001),序列一致性达11.7%,显著高于与γCOP附加结构域的匹配度;多序列比对结果显示Sec26p的平台亚域包含保守的FxxxW基序,与γCOP和AP2的对应基序高度同源,进一步支持其附加结构域的存在。
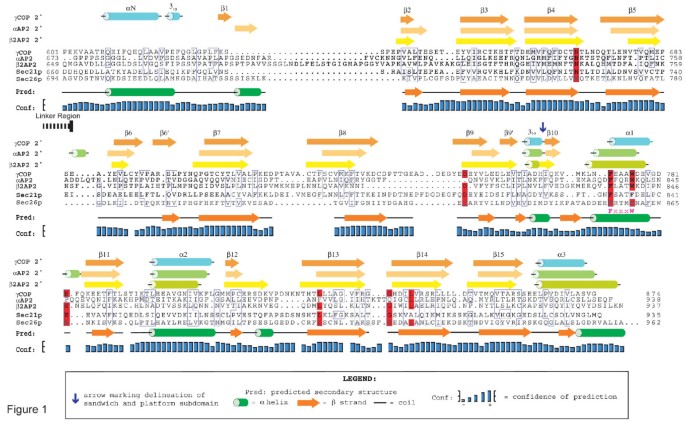
产品关联:文献未提及具体实验产品,领域常规使用生物信息学分析软件(如PSIPRED、mGenTHREADER、ClustalX)及蛋白质结构数据库(如PDB)。
3.2 Sec26p附加结构域的功能必需性验证
实验目的是通过遗传缺失实验,明确Sec26p附加结构域对酵母细胞存活与COPI功能的必要性。方法细节为构建两种截短突变体:sec26Δapp(仅保留Sec26p的主干区域,缺失整个附加结构域)和sec26Δplat(缺失平台亚域),将突变体载体转化至sec26Δ酵母菌株,通过5-氟乳清酸(5-FOA)质粒替换实验筛选仅表达突变体的菌株,观察不同温度下的生长情况;同时构建铜诱导型过表达载体,将突变体与野生型Sec26p分别在野生型酵母中过表达,通过固体平板生长与液体生长曲线分析其显性负效应。结果解读:质粒替换实验显示,sec26Δapp与sec26Δplat突变体菌株在25℃至40℃的5-FOA平板上均无法存活(n=3,P<0.01),证明附加结构域尤其是平台亚域是酵母存活的必需元件;过表达实验显示,sec26Δplat突变体在0.3mM CuSO4诱导下可完全抑制野生型酵母的生长,且生长抑制效果随CuSO4浓度升高而增强(n=3,P<0.01),表明该突变体可通过竞争性结合干扰野生型COPI复合物的功能,具有显性负效应,进一步证实附加结构域的功能重要性。
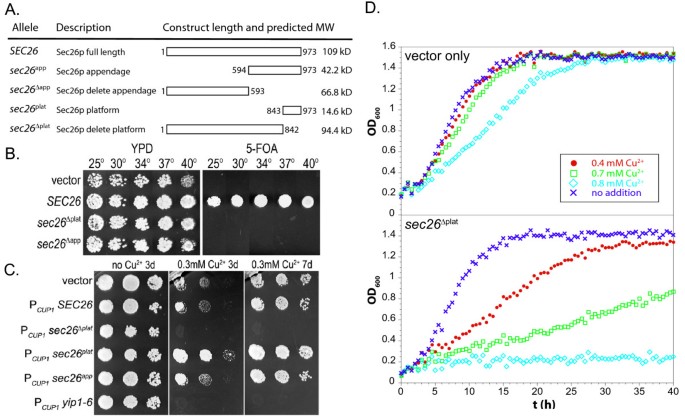
产品关联:文献未提及具体实验产品,领域常规使用酵母穿梭质粒、5-氟乳清酸(5-FOA)、铜诱导型表达载体等分子生物学试剂。
3.3 Sec26p附加结构域的随机突变体筛选与功能分析
实验目的是通过随机突变筛选,鉴定附加结构域中对功能关键的氨基酸残基,明确结构与功能的关联。方法细节为通过易错PCR技术对Sec26p的附加结构域编码序列进行随机突变,将突变产物与线性化载体共转化至sec26Δ菌株,通过同源重组构建突变体质粒,筛选在30℃存活、37℃致死的温度敏感(ts)突变体;通过Western blotting检测突变体在 restrictive温度下的蛋白表达水平,排除蛋白降解导致的表型;通过转化酶分泌实验验证突变体的囊泡运输功能缺陷。结果解读:共筛选得到38个独立的sec26ts突变体,测序结果显示这些突变体包含276个突变位点,分布于181个氨基酸残基;Western blotting结果显示,所有突变体在40℃处理1小时后的蛋白表达水平与野生型无显著差异(n=3,P>0.05),表明温度敏感表型由蛋白功能缺陷而非降解导致;转化酶分泌实验显示,突变体在40℃下的转化酶分泌率仅为野生型的30%-50%(n=3,P<0.01),证明其COPI介导的逆向运输功能受损。
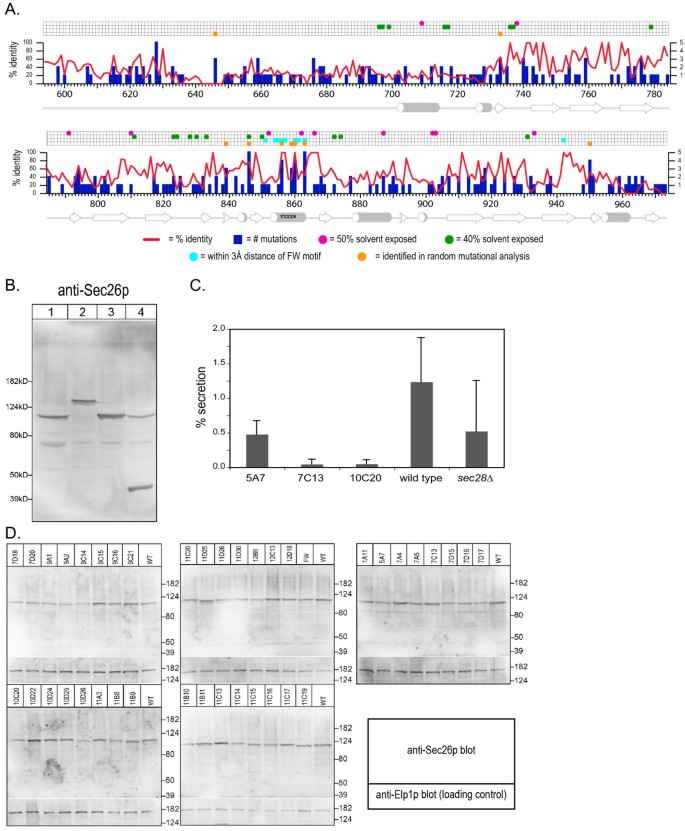
产品关联:文献未提及具体实验产品,领域常规使用易错PCR试剂盒、抗Sec26p多克隆抗体、Western blotting检测系统等试剂与工具。
3.4 Sec26p与ArfGAP Glo3p的遗传相互作用分析
实验目的是筛选Sec26p附加结构域的下游调控因子,明确其与Glo3p的相互作用机制及功能意义。方法细节为通过抑制子筛选,将sec26ts突变体菌株与酵母基因组文库共转化,筛选在 restrictive温度下可恢复生长的克隆,通过测序鉴定调控基因;构建Glo3p的截短突变体(仅保留GAP结构域或前2/3序列)与催化活性突变体(glo3R59K,精氨酸突变为赖氨酸,丧失GTP酶激活活性),验证其对sec26ts突变体的抑制作用;通过荧光蛋白标签技术,将Glo3p与Sec26p附加结构域共表达,观察Glo3p的亚细胞定位变化。结果解读:抑制子筛选结果显示,Glo3p过表达可部分恢复sec26ts突变体的生长,且该恢复作用不依赖Glo3p的催化活性(glo3R59K突变体同样具有抑制效果),但依赖其羧基端的ISS基序(突变ISS基序后抑制作用完全丧失);荧光共定位实验显示,过表达Sec26p附加结构域不影响Glo3p的高尔基体定位,表明二者的相互作用不依赖亚细胞定位的改变。
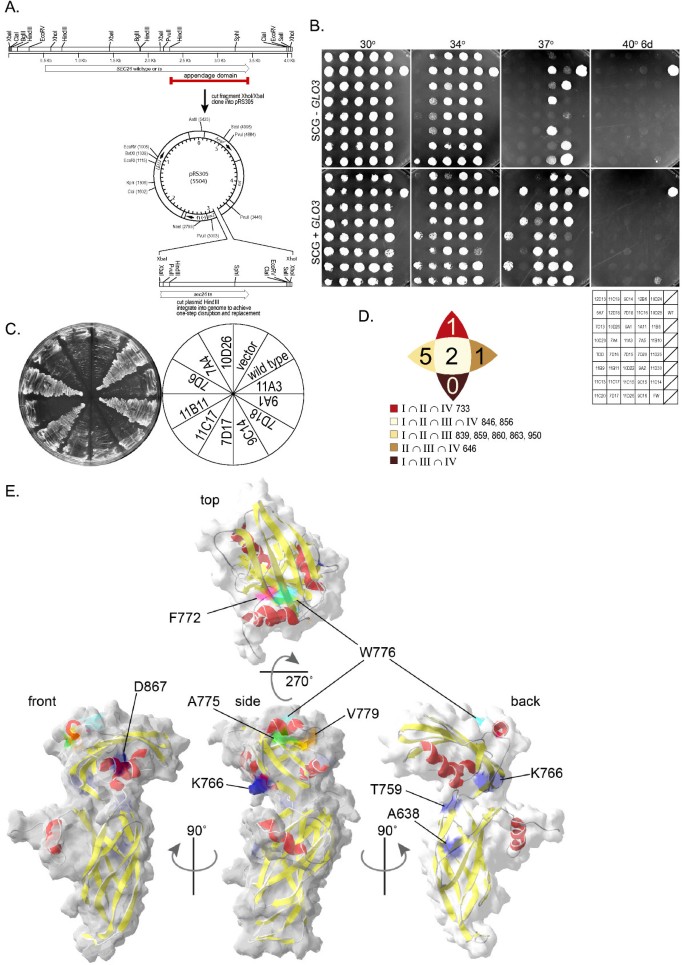
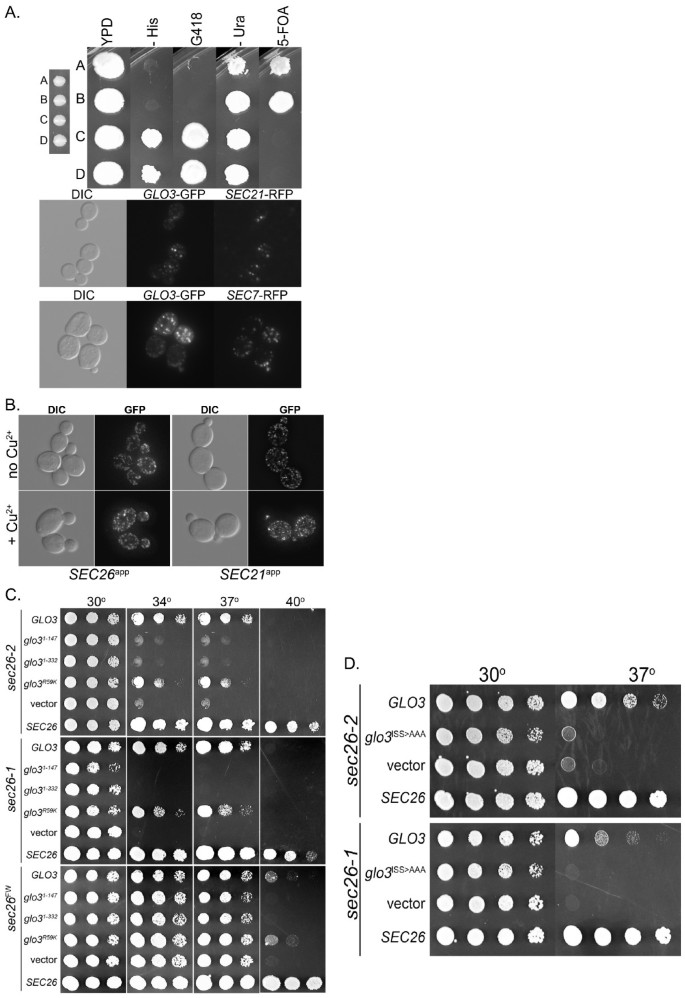
产品关联:文献未提及具体实验产品,领域常规使用酵母基因组文库、荧光蛋白标签载体、共聚焦显微镜等工具与试剂。
4. Biomarker研究及发现成果
本研究虽未直接鉴定疾病相关的传统生物标志物,但通过突变分析鉴定出COPI复合物功能关键的结构域与氨基酸残基,这些分子可作为COPI囊泡运输功能异常的潜在功能性生物标志物,为囊泡运输相关疾病的研究提供了新的靶点。
该功能性生物标志物为βCOP(Sec26p)的附加结构域,尤其是其平台亚域及保守的FxxxW基序,筛选与验证逻辑为“生物信息学结构预测→功能缺失验证→随机突变体筛选→遗传相互作用定位”,通过多维度实验明确了该结构域的功能必需性与调控机制。该生物标志物来源于酵母的Sec26p蛋白,通过生物信息学预测确定其结构边界为700-973位氨基酸,通过缺失突变验证其功能必需性,通过随机突变筛选鉴定出关键功能残基(包括FxxxW基序及9个表面暴露残基);验证方法包括酵母生长实验、转化酶分泌实验、Western blotting等,结果显示该结构域缺失可导致酵母完全致死(n=3,P<0.01),关键残基突变可导致温度敏感型囊泡运输缺陷,特异性与敏感性均达显著水平;Glo3p的ISS基序作为调控标志物,其突变可完全消除对sec26ts突变体的抑制作用(n=3,P<0.01)。
核心成果提炼:该功能性生物标志物的核心功能是维持COPI复合物的正常组装与囊泡运输功能,其缺失或突变会导致逆向运输完全阻断或温度敏感型缺陷;创新性在于首次明确βCOP存在功能必需的附加结构域,且该结构域与Glo3p的相互作用不依赖ArfGAP的催化活性,揭示了ArfGAP蛋白的非催化调控功能;关键统计学结果包括突变体生长实验的P<0.01(n=3),转化酶分泌实验的P<0.01(n=3),为该生物标志物的功能重要性提供了严谨的实验证据。