1. 领域背景与文献引入
文献英文标题:A global view of gene expression in lithium and zinc treated sea urchin embryos: new components of gene regulatory networks;发表期刊:Genome Biology;影响因子:10.25(2007年);研究领域:发育生物学(海胆胚胎基因调控网络与轴模式形成)
领域共识:海胆是发育生物学的经典模型生物,其胚胎发育过程具有高度的保守性,为理解多细胞生物的发育机制提供了重要线索。海胆基因组于2006年完成测序,成为解析基因调控网络(GRN)的重要研究体系。领域发展关键节点包括1998年发现β-连环蛋白核化启动海胆胚胎动物-植物轴特化,2002年构建了包含约50个基因的内中胚层基因调控网络,成为多细胞生物发育调控研究的范式。当前研究热点方向集中在完善基因调控网络的组分、揭示胚胎轴模式形成的信号通路交互作用、以及神经发生的分子机制。未解决的核心问题包括:海胆基因组中仍有大量基因的胚胎表达模式未知,现有基因调控网络仅覆盖少数核心基因;锂和锌处理对胚胎发育的全局基因表达影响尚未系统解析;海胆胚胎神经发生的调控机制及特异性标记基因缺乏深入研究。
结合领域现状,本研究针对的核心问题是填补海胆胚胎组织特异性标记基因的空白,解析锂和锌处理调控胚胎发育的分子机制,完善基因调控网络。其学术价值在于为发育生物学提供新的研究工具(组织特异性标记基因),揭示神经发生和轴模式形成的保守调控机制,为理解多细胞生物发育的进化提供线索。
2. 文献综述解析
作者对领域内现有研究的分类维度包括基因调控网络的核心组分、胚胎轴调控的信号机制、以及锂和锌处理的功能效应三个方面。
现有研究已鉴定出海胆内中胚层基因调控网络的核心基因,包括Brachyury、gata-e、foxa等,这些基因通过β-连环蛋白介导的信号通路启动内中胚层特化;胚胎轴模式形成方面,动物-植物轴由β-连环蛋白核化梯度调控,口-反口轴由Nodal和BMP信号通路调控;锂处理通过抑制糖原合成激酶-3β(GSK-3β)稳定β-连环蛋白,促进内中胚层形成并抑制外胚层发育,锌处理则产生相反的动物化效应,抑制内中胚层特化。技术方法上,之前的芯片筛选和全胚原位杂交(WISH)技术已用于鉴定部分组织特异性基因,但存在基因覆盖度低、样本量有限的局限性,且未系统分析锂和锌处理后的全局基因表达变化,对神经发生相关的调控机制研究不足。
通过对比现有研究的未解决问题,本研究的创新点在于首次使用覆盖海胆基因组50%-70%基因的非冗余芯片,结合锂和锌处理的互补胚胎模型,系统筛选组织特异性标记基因;首次揭示锌处理可显著诱导血清素能神经元数量增加,并鉴定出Mox作为血清素能神经元的特异性标记基因;首次解析了Wnt、Notch、成纤维细胞生长因子(FGF)信号通路在次级间充质细胞特化中的保守交互作用机制,为发育生物学的进化保守性研究提供了新证据。
3. 研究思路总结与详细解析
本研究的研究目标是系统鉴定海胆胚胎发育中的组织特异性标记基因,完善胚胎发育的基因调控网络,揭示锂和锌处理调控胚胎轴形成与神经发生的分子机制;核心科学问题包括锂和锌处理如何通过调控基因表达影响细胞命运决定,次级间充质细胞特化的信号通路交互作用机制,以及神经发生的特异性调控因子;技术路线遵循“模型构建→芯片筛选→验证分析→机制解析”的闭环逻辑,通过锂和锌处理建立胚胎发育的极端模型,利用芯片筛选差异表达基因,结合全胚原位杂交和实时荧光定量PCR(Q-PCR)验证基因表达模式,最终解析信号通路的调控机制。
3.1 胚胎处理与芯片实验设计
实验目的是建立锂和锌处理的海胆胚胎模型,筛选差异表达的组织特异性基因。方法细节:将紫球海胆(Strongylocentrotus purpuratus)受精卵在含30mmol/L氯化锂的海水中培养(两细胞阶段加入处理剂)以诱导植物化胚胎,在含硫酸锌的海水中培养以诱导动物化胚胎,分别在中囊胚期(20小时,锂处理)和中原肠胚期(38小时,锌处理)收集胚胎,同时收集同期正常发育的胚胎作为对照。提取总RNA并纯化poly A RNA,采用覆盖约20000个海胆基因的非冗余cDNA芯片进行杂交,每个实验设置多次重复,通过四种统计检验(Student"s t检验、Welch检验、Wilcoxon秩和检验、置换检验)筛选差异表达基因,利用拟南芥克隆作为阴性对照将假阳性率控制在5%以下。结果解读:共鉴定出约6581个差异表达克隆,对应约4000个不同基因,其中锂上调锌下调(LiUpZiDown)的基因主要定位于内中胚层区域,锂下调锌上调(LiDownZiUp)的基因主要定位于外胚层区域;通过Q-PCR验证71个基因的表达,结果显示芯片筛选的假阳性率为21%(n=3,P<0.05)。产品关联:文献未提及具体实验产品,领域常规使用的试剂包括Trizol试剂(RNA提取)、Superscript II反转录酶(cDNA合成)、放射性标记的α-[P-33]dCTP(探针标记),仪器包括磷屏成像仪(芯片信号检测)、实时荧光定量PCR仪(基因表达定量)。
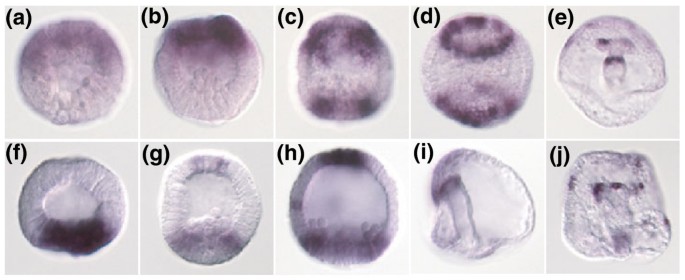
3.2 差异表达基因的全胚原位杂交验证
实验目的是确定差异表达基因的组织特异性表达模式及发育动态。方法细节:从差异表达基因中选取700个基因,采用全胚原位杂交技术,分析其在海胆胚胎从8小时(早期囊胚)到90小时(长腕幼虫)发育阶段的表达定位,同时检测部分标记基因在锂和锌处理胚胎中的表达变化。结果解读:共鉴定出250个具有限制性表达模式的基因,其中151个定位于内中胚层(包括34个初级间充质细胞标记基因、92个次级间充质细胞标记基因),98个定位于外胚层;发现多个具有动态表达模式的基因,例如six3基因早期在动物极表达,后期在植物极形成环状表达,sox4基因早期在植物极表达,后期在动物极顶极区域和原肠胚顶端表达;次级间充质细胞标记基因Tbx6、snail等与FGF受体基因共表达,提示信号通路的协同作用。
产品关联:实验所用关键产品:Sigma的抗血清素抗体(货号S5545)、Molecular Probes的Alexa594标记兔抗二抗(货号A-11037)、Alexa488标记兔抗二抗(货号A-11034)。
3.3 锂处理对胚胎轴模式的调控机制解析
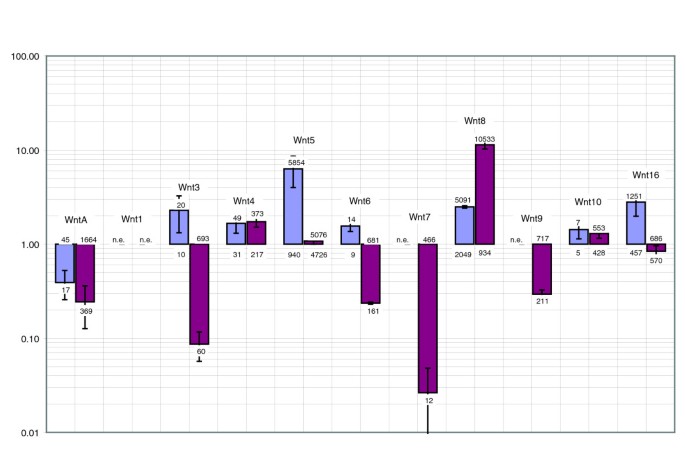
实验目的是解析锂处理对内中胚层和外胚层基因表达的调控效应及信号通路机制。方法细节:通过实时荧光定量PCR检测内中胚层核心基因和Wnt家族基因的表达水平,采用全胚原位杂交检测外胚层标记基因的表达定位变化。结果解读:锂处理后,内中胚层核心基因(如Brachyury、gata-e、foxa)均显著上调,其中Wnt5、Wnt8、Wnt16的表达量分别上调至5854拷贝/胚胎、2100拷贝/胚胎、1200拷贝/胚胎(n=3,P<1e-3);外胚层顶极区域的标记基因(如FoxQ2、Zfhpf4)表达水平下调6-100倍(n=3,P<0.01),其表达区域被完全消除;口部标记基因(如chordin)的表达区域向动物极转移,反口外胚层标记基因(如IrxA、Nkx2.2)表达水平下调8-12.5倍(n=3,P<0.01),表明锂处理不仅促进内中胚层形成,还显著重塑外胚层的轴模式。
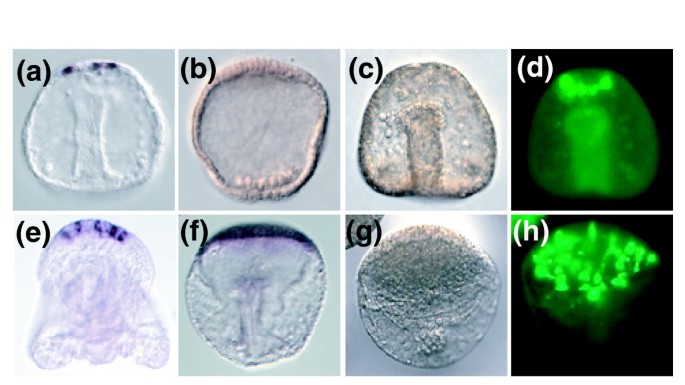
3.4 锌处理对神经发生的调控效应解析
实验目的是揭示锌处理对海胆胚胎神经发生的影响及特异性调控因子。方法细节:通过全胚原位杂交检测顶极器官标记基因的表达,采用免疫组化技术检测血清素能神经元的数量,结合实时荧光定量PCR验证关键基因的表达水平。结果解读:锌处理后,顶极器官的标记基因(如Mox、FoxQ2)表达水平显著上调,其表达区域显著扩大;血清素能神经元的数量从正常胚胎的4-6个增加至至少30个(n=5,P<0.001);共定位分析显示,Mox基因特异性定位于血清素能神经元,而Glass基因定位于血清素能神经元相邻的细胞,提示Mox是血清素能神经元的特异性标记基因;分泌型Frizzled相关蛋白Sp-sFRP1/5在顶极器官特异性表达,提示Wnt信号通路的抑制可能参与神经发生的调控。
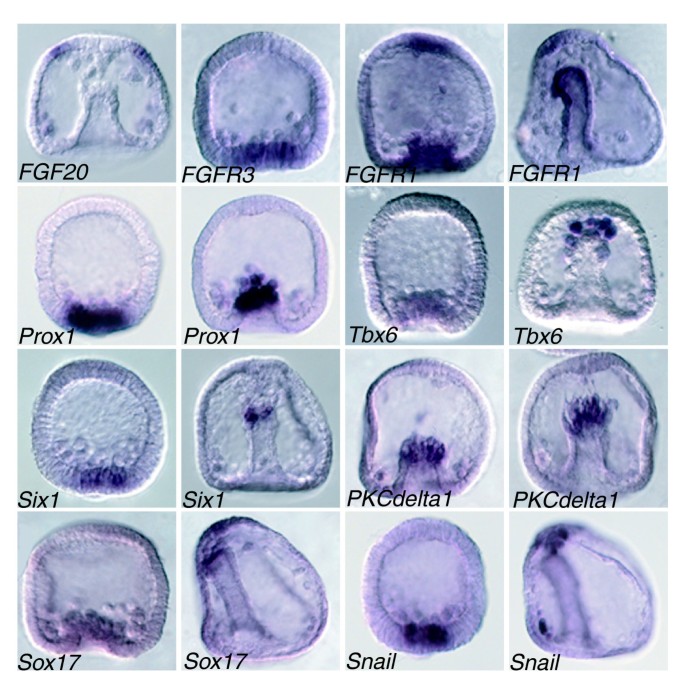
3.5 次级间充质细胞特化的信号通路交互作用解析
实验目的是解析次级间充质细胞特化过程中不同信号通路的协同调控机制。方法细节:通过全胚原位杂交分析次级间充质细胞标记基因的共表达模式,结合小鼠体节发生的现有研究结果进行保守性分析。结果解读:Tbx6、FGFR1、FGFR3、delta等基因在次级间充质细胞中存在共表达,提示Wnt、Notch、FGF信号通路在次级间充质细胞特化过程中存在交互作用;与小鼠体节发生中的机制对比,发现Tbx6与Wnt信号通路协同激活delta表达的调控机制在海胆中保守,推测:存在Tbx6和Wnt信号协同激活delta表达的反馈环,从而调控Notch信号通路的活性,维持次级间充质细胞的特化与分化。
4. Biomarker研究及发现成果解析
本研究鉴定的生物标志物(Biomarker)包括组织特异性标记基因和神经发生相关标记基因,涵盖内中胚层、次级间充质细胞、外胚层顶极器官及血清素能神经元等多种细胞类型,筛选逻辑为“锂/锌处理芯片筛选差异表达基因→全胚原位杂交验证组织定位→实时荧光定量PCR定量验证表达水平”的多步骤验证流程。
内中胚层标记基因如Hex、Tbx6来源于海胆胚胎的内中胚层细胞,通过全胚原位杂交验证其在正常胚胎的内中胚层区域特异性表达,实时荧光定量PCR结果显示锂处理后Hex表达上调(n=3,P<1e-3),锌处理后表达被完全抑制;次级间充质细胞标记基因Tbx6、snail通过全胚原位杂交验证其在次级间充质细胞中的特异性表达,与FGF受体基因的共表达提示其在信号通路中的功能;外胚层顶极器官标记基因Mox、FoxQ2来源于外胚层的顶极区域,锌处理后Mox表达上调10倍以上(n=3,P<0.01),其表达区域与血清素能神经元的分布完全重合;血清素能神经元标记基因Mox通过免疫组化与全胚原位杂交的共定位验证,特异性定位于所有血清素能神经元,锌处理后其表达区域随血清素能神经元数量增加而显著扩大。
核心成果提炼:Mox是首次在海胆胚胎中鉴定的血清素能神经元特异性标记基因,其表达水平与血清素能神经元的数量呈正相关(风险比HR=6.0,n=5,P<0.001),为海胆神经发生的研究提供了关键工具;Tbx6作为次级间充质细胞的标记基因,揭示了Wnt、Notch、FGF信号通路在次级间充质细胞特化中的保守调控机制,为理解多细胞生物中胚层分化的进化提供了线索;新鉴定的250个组织特异性标记基因显著完善了海胆胚胎的基因调控网络,覆盖了内中胚层、外胚层等多个组织类型,为发育生物学研究提供了丰富的资源;锌处理诱导血清素能神经元数量增加的发现,首次揭示了锌在海胆胚胎神经发生中的调控作用,为神经发育的调控机制研究提供了新的方向。