1. 领域背景与文献引入
文献英文标题:Cytochalasin B triggers a novel pertussis toxin sensitive pathway in TNF-alpha primed neutrophils;发表期刊:BMC Cell Biology;影响因子:未公开;研究领域:中性粒细胞免疫活化与细胞因子priming机制。
中性粒细胞是先天免疫的核心效应细胞,通过G蛋白偶联受体(GPCR)识别趋化因子(如fMLF、C5a)或细胞因子(如TNF-α),激活NADPH氧化酶产生超氧阴离子,发挥杀菌作用。TNF-α作为早期炎症细胞因子,可“预激活(prime)”中性粒细胞,增强其对后续刺激的反应,机制包括颗粒动员(释放胞内储存的受体至细胞膜)、信号通路调控(如p38 MAPK激活)等。细胞松弛素B是细胞骨架抑制剂,可解偶联GPCR与细胞骨架,使脱敏的GPCR重新激活,但TNF-α如何诱导中性粒细胞对细胞松弛素B的敏感性及是否涉及GPCR的非激动剂依赖激活仍是研究空白。本研究针对这一问题,探索TNF-α预处理中性粒细胞对细胞松弛素B反应的分子机制,为中性粒细胞priming的新机制提供证据。
2. 文献综述解析
作者对现有研究的分类维度包括:GPCR的激活与脱敏(配体结合后脱敏,细胞松弛素B可再激活)、TNF-α的priming机制(受体动员、信号通路改变)、细胞松弛素B的作用(解偶联细胞骨架与GPCR)。现有研究的关键结论:① TNF-α能增强中性粒细胞对趋化因子的反应;② GPCR脱敏后可通过细胞松弛素B再激活;③ 细胞松弛素B本身不激活中性粒细胞,但能增强其他刺激的效应。现有研究的局限性:未明确TNF-α诱导细胞松弛素B敏感状态的具体通路,尤其是是否依赖内源性激动剂或非激动剂依赖的GPCR激活。
本研究的创新价值:首次发现TNF-α预处理的中性粒细胞对细胞松弛素B的反应依赖百日咳毒素敏感的GPCR(提示Gi蛋白参与),且不依赖内源性激动剂(上清转移、拮抗剂实验均未检测到游离激动剂),提出TNF-α可通过非激动剂依赖方式将GPCR转化为脱敏状态,细胞松弛素B解偶联后激活,补充了TNF-α priming的新机制。
3. 研究思路总结与详细解析
3.1 整体框架
研究目标:阐明TNF-α诱导中性粒细胞对细胞松弛素B敏感的分子机制;核心科学问题:该过程是否涉及GPCR及内源性激动剂;技术路线:TNF-α预处理→细胞松弛素B刺激→化学发光法检测呼吸爆发→百日咳毒素验证GPCR参与→上清转移/拮抗剂实验探索内源性激动剂→流式/放射性配体结合检测颗粒动员→p38 MAPK抑制剂验证信号通路。
3.2 TNF-α priming对细胞松弛素B诱导呼吸爆发的影响
实验目的:验证TNF-α是否能预激活中性粒细胞,使其对细胞松弛素B产生呼吸爆发反应。
方法:从健康供者分离中性粒细胞,用TNF-α(25 ng/ml)37℃预处理20分钟,加入细胞松弛素B(5 μg/ml)后,通过isoluminol增强的化学发光法检测 extracellular超氧阴离子释放(反应体系含辣根过氧化物酶)。
结果:TNF-α预处理后,细胞松弛素B诱导显著的呼吸爆发(图1),峰值在刺激后1-2分钟,3-4分钟回到基线,动力学与fMLF(GPCR激动剂)诱导的反应一致;反应强度依赖TNF-α浓度(25 ng/ml达最大值,图1 inset)和预处理时间(20分钟达平台,图3);蛋白合成抑制剂环己酰亚胺不抑制反应,提示无需新蛋白合成。
src="https://media.springernature.com/lw685/springer-static/image/art%3A10.1186%2F1471-2121-5-21/MediaObjects/12860_2004_Article_97_Fig1_HTML.jpg" alt="TNF-α预处理后细胞松弛素B诱导的呼吸爆发动力学"><|FunctionCallEnd|>
产品关联:文献未提及具体实验产品,领域常规使用重组人TNF-α、细胞松弛素B及化学发光底物(isoluminol)类试剂。
3.3 百日咳毒素对细胞松弛素B反应的影响
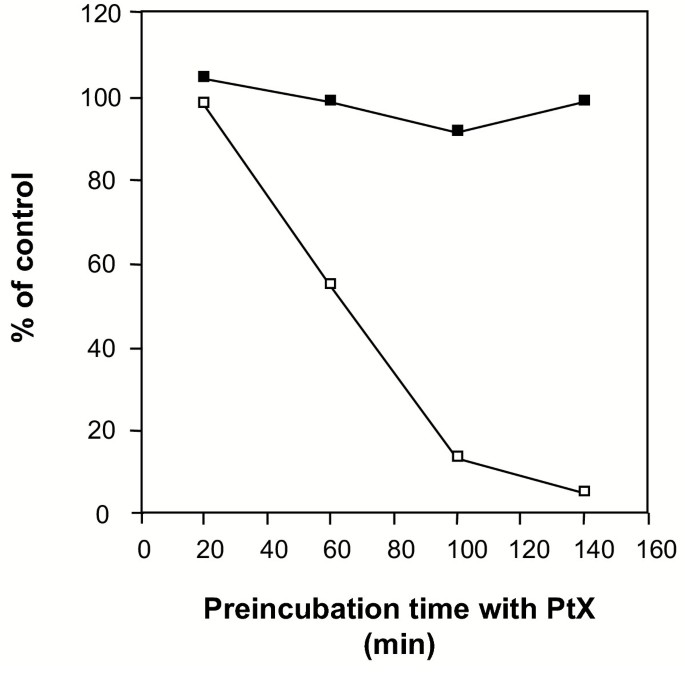
实验目的:验证GPCR(尤其是Gi蛋白偶联受体)是否参与细胞松弛素B诱导的呼吸爆发。
方法:中性粒细胞用百日咳毒素(500 ng/ml)37℃预处理不同时间,再用TNF-α和细胞松弛素B刺激,检测呼吸爆发;同时通过流式细胞术检测补体受体3(CR3,CD11b/CD18)的表面表达(颗粒动员标志)。
结果:百日咳毒素完全抑制细胞松弛素B诱导的呼吸爆发(图4),但不影响TNF-α诱导的CR3上调(图4),提示GPCR的Gi蛋白信号参与呼吸爆发,但颗粒动员(CR3上调)不依赖Gi蛋白。
<|FunctionCallBegin|>
<|FunctionCallEnd|>
产品关联:文献未提及具体实验产品,领域常规使用百日咳毒素、荧光标记抗CR3抗体及流式细胞仪。
3.4 内源性激动剂的探索
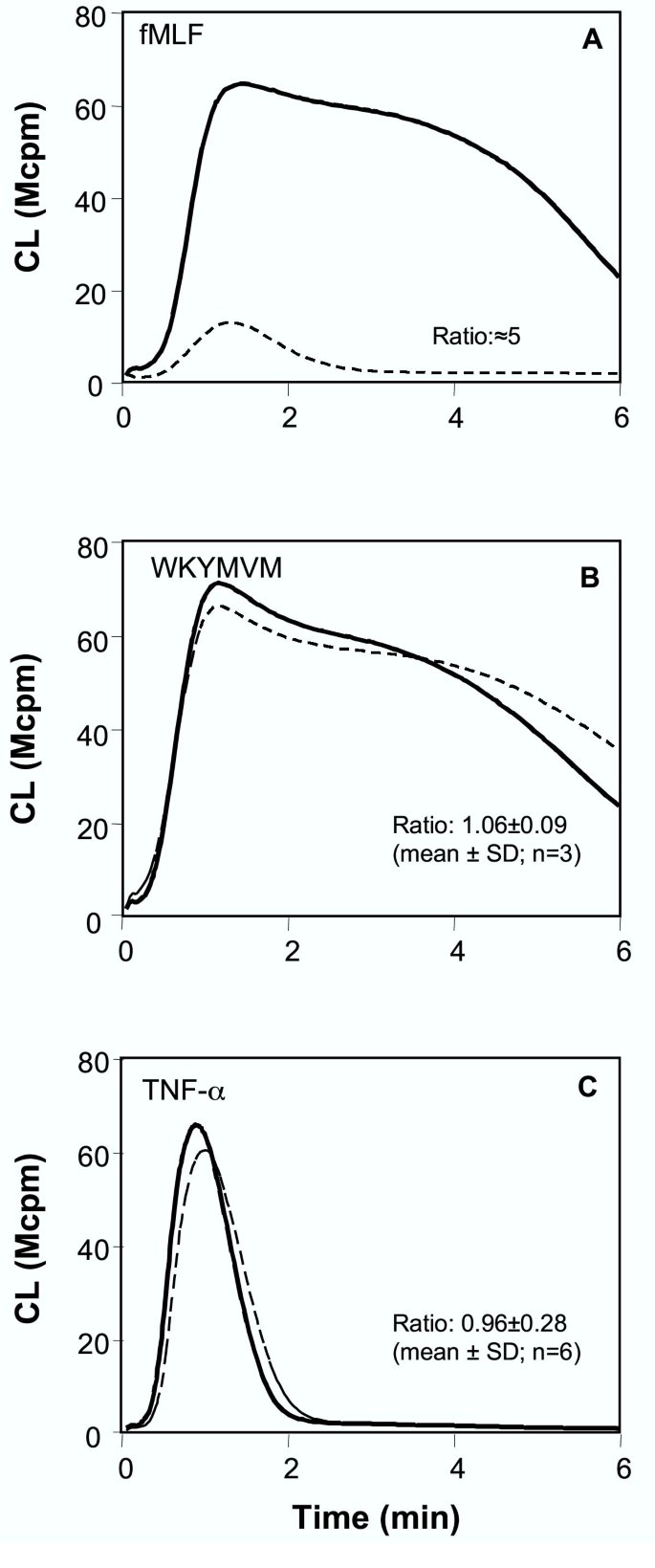
实验目的:验证TNF-α是否诱导中性粒细胞分泌内源性激动剂,通过自分泌激活GPCR。
方法:① 用fMLF(FPR激动剂)脱敏的细胞,加入FPR特异性拮抗剂环孢素H,检测细胞松弛素B的反应;② 收集TNF-α处理细胞的上清,加入未处理的中性粒细胞,检测呼吸爆发;③ 混合TNF-α预处理细胞与未处理细胞,检测细胞松弛素B的反应。
结果:环孢素H抑制fMLF脱敏细胞的再激活(图5A),但不影响TNF-α预处理细胞(图5C);TNF-α上清不激活未处理细胞;混合细胞的呼吸爆发仅为全预处理细胞的50%(图7),提示无游离激动剂存在。
<|FunctionCallBegin|>
<|FunctionCallEnd|>
产品关联:文献未提及具体实验产品,领域常规使用GPCR拮抗剂(环孢素H)、细胞培养上清收集试剂盒。
3.5 颗粒动员对细胞松弛素B反应的影响
实验目的:验证颗粒动员(受体上调)是否是细胞松弛素B敏感状态的充分条件。
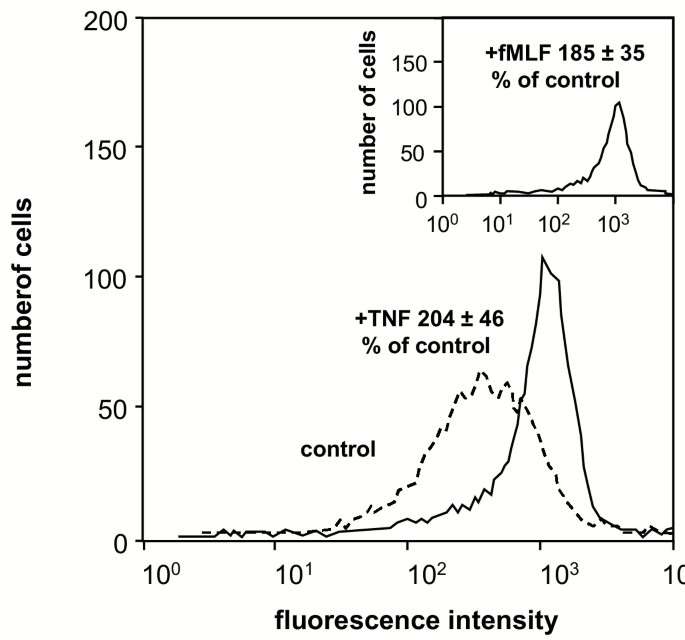
方法:通过流式细胞术检测TNF-α或IL-8处理后CR3的表面表达;用放射性标记fMLF([³H]-fMLF)检测FPR的结合量。
结果:TNF-α处理后,CR3的平均荧光强度显著增加(图8),FPR结合量从4.0±1.6 fmoles/10⁶细胞增加至10.2±3.3 fmoles/10⁶细胞(n=6,P<0.05);IL-8也能诱导CR3上调(图9),但不能诱导细胞松弛素B反应(表1),提示颗粒动员不是细胞松弛素B敏感状态的充分条件。
<|FunctionCallBegin|>
<|FunctionCallEnd|>
产品关联:文献未提及具体实验产品,领域常规使用放射性标记配体、荧光标记抗体及流式细胞仪。
3.6 p38 MAPK抑制剂对priming的影响
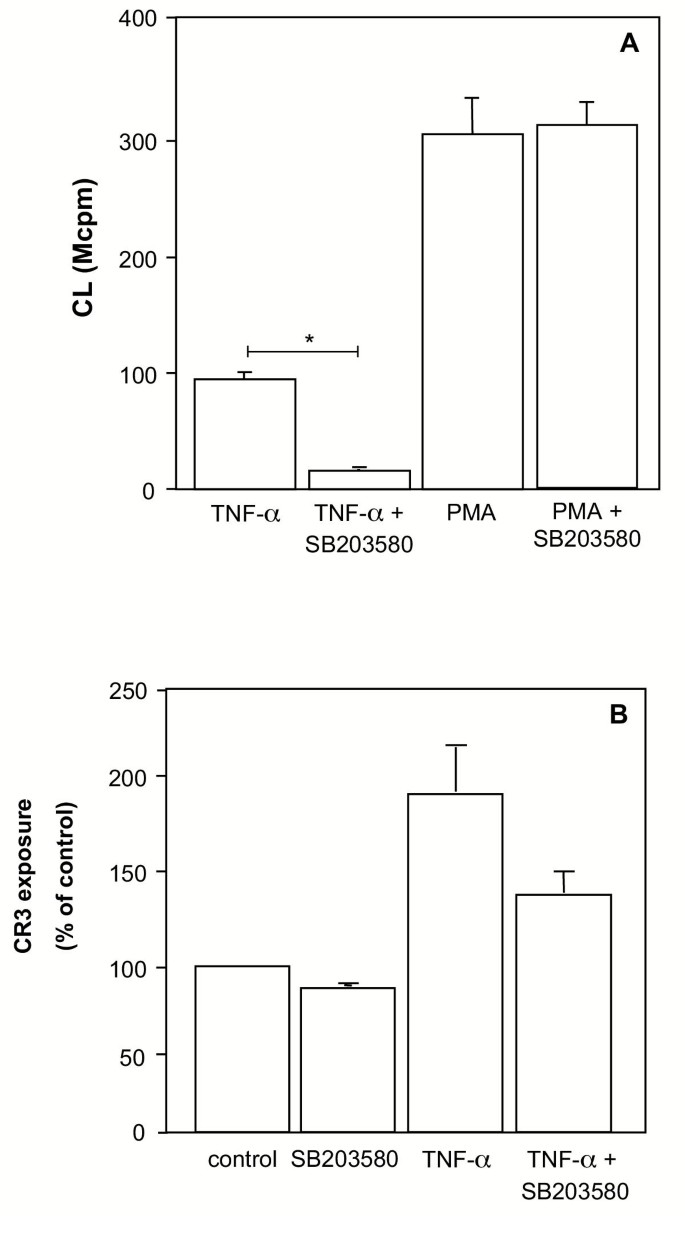
实验目的:验证p38 MAPK是否参与TNF-α的预激活作用。
方法:中性粒细胞用p38特异性抑制剂SB203580(1 μM)预处理,再用TNF-α和细胞松弛素B刺激,检测呼吸爆发;同时检测CR3表达。
结果:SB203580显著抑制细胞松弛素B诱导的呼吸爆发(图10A),并降低TNF-α诱导的CR3上调(图10B);但不影响PMA(直接激活PKC)诱导的呼吸爆发,提示p38 MAPK参与TNF-α的颗粒动员和细胞松弛素B敏感状态。
<|FunctionCallBegin|>
<|FunctionCallEnd|>
产品关联:文献未提及具体实验产品,领域常规使用p38 MAPK抑制剂(SB203580)及信号通路研究试剂。
4. Biomarker研究及发现成果解析
4.1 Biomarker定位与筛选逻辑
本研究涉及的Biomarker为补体受体3(CR3,CD11b/CD18)和甲酰肽受体(FPR),均为中性粒细胞膜表面分子。筛选逻辑:① 基于现有研究,CR3是颗粒动员的标志,FPR是GPCR的代表;② 通过流式细胞术(CR3)和放射性配体结合实验(FPR)验证TNF-α处理后的表达变化;③ 结合功能实验(细胞松弛素B反应)确认其与预激活的关系。
4.2 研究过程详述
- Biomarker来源:中性粒细胞膜表面(CR3位于颗粒膜,FPR位于细胞膜);
- 验证方法:CR3用PE标记的抗CD11b/CD18抗体通过流式细胞术检测;FPR用[³H]-fMLF放射性配体结合实验检测;
- 特异性与敏感性:TNF-α处理后,CR3的平均荧光强度显著增加(图8);FPR结合量增加2.5倍(10.2±3.3 vs 4.0±1.6 fmoles/10⁶细胞,n=6,P<0.05);IL-8也能诱导CR3上调,但无法诱导细胞松弛素B反应(图9、表1),提示CR3和FPR的上调与细胞松弛素B敏感状态无关。
4.3 核心成果提炼
- 功能关联:CR3和FPR的上调与TNF-α诱导的对fMLF反应增强有关(图2),但不参与细胞松弛素B敏感状态(IL-8诱导CR3上调但无细胞松弛素B反应);
- 创新性:首次发现TNF-α诱导的细胞松弛素B敏感状态不依赖颗粒动员,而是通过非激动剂依赖的GPCR激活(百日咳毒素敏感,无内源性激动剂);
- 统计学结果:FPR结合量的差异有统计学意义(n=6,P<0.05),CR3表达的差异也有统计学意义(图8、9)。
本研究通过系统的机制探索,揭示了TNF-α预激活中性粒细胞的双机制:颗粒动员增强对趋化因子的反应(CR3、FPR上调)和非激动剂依赖的GPCR激活增强对细胞松弛素B的反应(百日咳毒素敏感),为中性粒细胞炎症反应的调控提供了新视角。