1. 领域背景与文献引入
文献英文标题:Sex chromosome-to-autosome transposition events counter Y-chromosome gene loss in mammals;发表期刊:Genome Biology;影响因子:未公开;研究领域:哺乳动物性染色体进化。
哺乳动物X和Y染色体起源于一对共同的常染色体祖先,但经过约1.6亿年的进化已高度分化:Y染色体尺寸仅为X的1/3~1/5,丢失了原始约640个基因中的95%,仅保留36个具有调控功能的特殊基因(涉及染色质修饰、转录、翻译等多个层面)。这些保留的Y连锁基因及其X连锁同源物具有更高的剂量敏感性,受自然选择维持两性中的双拷贝。然而,多数哺乳动物谱系仍经历了保守Y连锁基因的延迟丢失——例如日本刺鼠(Tokudaia osimensis)的Y染色体完全消失,其Y连锁基因通过转座到常染色体或X染色体得以保留,但这一现象此前被视为孤立案例。
性染色体向常染色体的转座事件此前被认为主要由减数分裂性染色体失活(MSCI)驱动(MSCI导致雄性减数分裂时性连锁基因沉默,迫使必需基因转座到常染色体),但对Y连锁基因丢失是否会触发类似转座补偿仍不清楚。本研究针对这一空白,系统分析了8种哺乳动物的基因组,发现Y连锁基因丢失的转座补偿机制广泛存在,刷新了对性染色体转座驱动因素的认知。
2. 文献综述解析
文献综述的核心评述逻辑围绕“Y染色体基因丢失的补偿机制”与“性染色体转座的驱动因素”展开:作者首先总结现有研究对Y染色体进化的共识——Y染色体基因丢失非随机,保留基因是具有调控功能的特殊集合;接着指出此前对补偿机制的局限——仅在日本刺鼠中发现转座补偿,被视为孤立案例;最后评述性染色体转座的驱动因素——现有研究主要聚焦MSCI,未考虑Y连锁基因丢失的作用。
现有研究的关键结论包括:① 哺乳动物Y染色体源于常染色体,但丢失大部分基因,保留的基因具有剂量敏感性;② 日本刺鼠的Y染色体消失后,Y连锁基因通过转座到常染色体获救;③ 性染色体转座主要由MSCI驱动(迫使睾丸必需基因转座)。现有研究的局限性在于:① 未认识到转座补偿机制的广泛性;② 对性染色体转座的驱动因素理解不全面。
本研究的创新价值在于:首次证明Y连锁基因丢失的转座补偿机制广泛存在于哺乳动物中(涉及有袋类、灵长类、啮齿类等多个谱系),且Y连锁基因丢失是性染色体转座的额外驱动因素——这补充了此前仅关注MSCI的理论框架,说明性染色体转座是“MSCI驱动+Y基因丢失驱动”的双重结果。
3. 研究思路总结与详细解析
3.1 整体框架
研究目标:验证哺乳动物中Y连锁基因丢失是否通过性染色体向常染色体转座实现补偿;核心科学问题:Y连锁基因丢失的转座补偿机制是否广泛存在;技术路线:基因组搜索转座基因→功能性验证(ORF+转录活性)→系统发育分析起源→表达模式分析功能的闭环。
3.2 目标基因筛选与基因组搜索
实验目的:从8种哺乳动物(人、黑猩猩、恒河猴、狨猴、小鼠、大鼠、牛、负鼠)的基因组中,筛选Y连锁基因丢失后转座到常染色体的同源物。
方法细节:聚焦7个满足“Y连锁基因丢失”且“单拷贝”的基因(AMELY、EIF1AY、EIF2S3Y、KDM5D、RPS4Y、UBA1Y、USP9Y),通过基因组比对搜索其常染色体同源物。
结果解读:发现4个基因(EIF1A、EIF2S3、RPS4、UBA1)在至少一个谱系中存在常染色体转座拷贝——例如啮齿类(小鼠、大鼠)丢失EIF1AY,但存在常染色体EIF1A拷贝;灵长类(人、恒河猴)丢失EIF2S3Y,但存在常染色体EIF2S3拷贝。
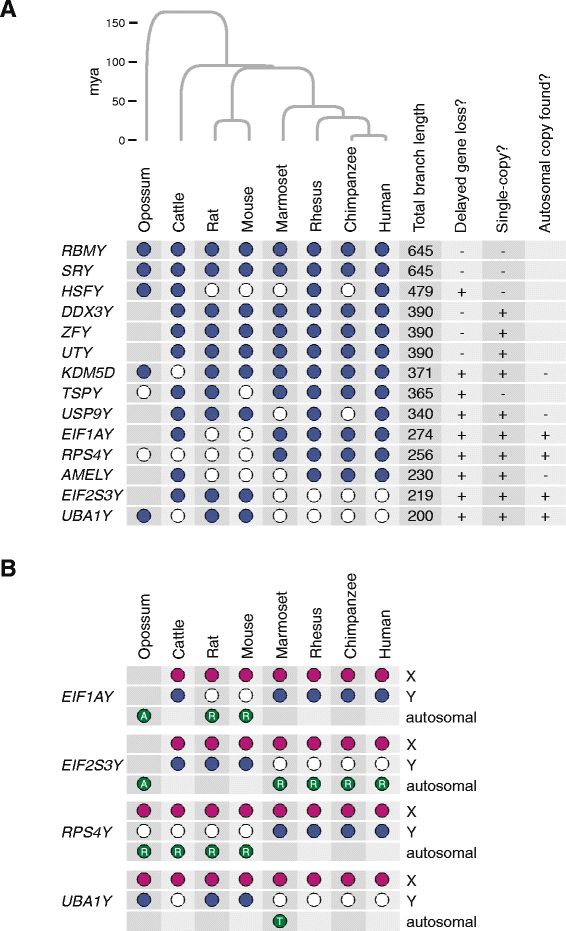
(Figure 1:展示8种哺乳动物Y连锁基因分布及常染色体同源物)
3.3 转座基因功能性验证
实验目的:确认转座到常染色体的基因是否为功能基因(非假基因)。
方法细节:① ORF完整性验证:将转座基因与性连锁同源物对比,检查是否存在截短或移码突变;② 转录活性验证:利用公共RNA-seq数据集(来自GenBank)检测转座基因的表达;③ 排除近期起源:若转座基因与亲本拷贝的核苷酸差异<2%,则视为近期起源(功能相关性低)。
结果解读:4个基因的常染色体转座拷贝均满足功能标准——例如小鼠EIF1A常染色体拷贝的ORF与X连锁同源物完全一致,且RNA-seq显示其在脑、肝等组织中高表达;灵长类EIF2S3常染色体拷贝虽位于不同基因组位置,但均有转录活性。
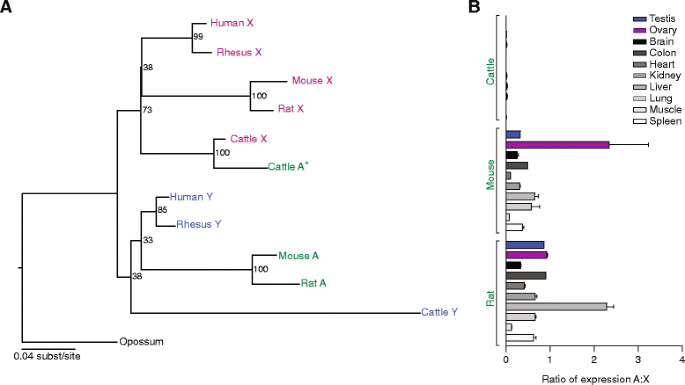
(Figure 2:展示EIF1A的系统发育与表达模式)
3.4 系统发育与选择压力分析
实验目的:解析转座事件的进化起源(亲本基因是X还是Y连锁)及选择压力(是否受净化选择)。
方法细节:① 序列比对:用MUSCLE对性连锁与常染色体转座基因的编码序列进行比对;② 系统发育树构建:用PhyML构建最大似然树(100次bootstrap验证);③ 选择压力计算:用PAL2NAL计算同义替换率(dS)与非同义替换率(dN)的比值(dN/dS)——dN/dS<1表明净化选择(功能保守)。
结果解读:转座事件为独立多次发生——例如EIF2S3在灵长类中独立转座3次(新世界猴、旧世界猴、猿类),系统发育树显示其亲本基因为X连锁;dN/dS比值均远低于1(如人-松鼠猴EIF2S3的dN/dS=0.075,小鼠-大鼠EIF1A的dN/dS=0.0010),表明转座基因受强净化选择,功能高度保守。
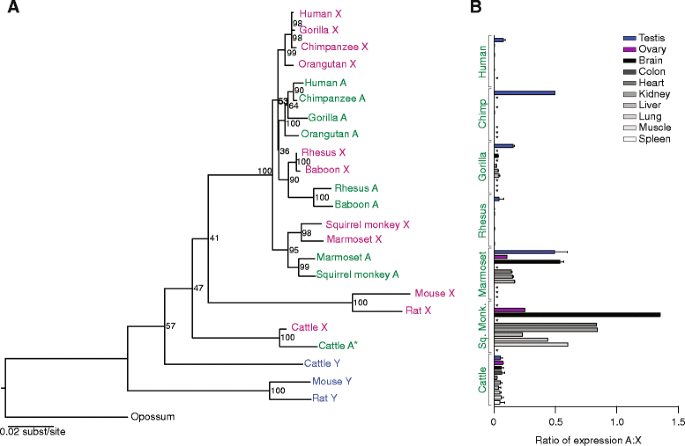
(Figure 3:展示EIF2S3的系统发育与表达模式)
3.5 表达模式分析
实验目的:探讨转座基因的功能分工(保留广泛表达或特化到睾丸)。
方法细节:下载GenBank中的RNA-seq数据集(覆盖脑、肝、肾、睾丸等多组织),用Bowtie将reads映射到常染色体与性连锁同源物,计算归一化后的表达量比值(常染色体/性连锁)。
结果解读:转座基因的表达模式分为两类——① 广泛表达:如啮齿动物的EIF1A常染色体拷贝在脑、肝、肾等多组织中表达,与X连锁拷贝的FPKM值相当(小鼠睾丸中FPKM=12.3 vs X连锁的11.8,n=3,P>0.05);② 睾丸特异性/优势表达:如人EIF2S3、牛RPS4的常染色体拷贝仅在睾丸中高表达(人睾丸中FPKM=25.6 vs X连锁的3.1,n=3,P<0.01),而其性连锁同源物在多组织中表达。这表明转座基因可通过两种方式补偿Y连锁基因丢失:要么保留广泛的剂量敏感性功能,要么特化维持雄性特异性功能(如精子发生)。
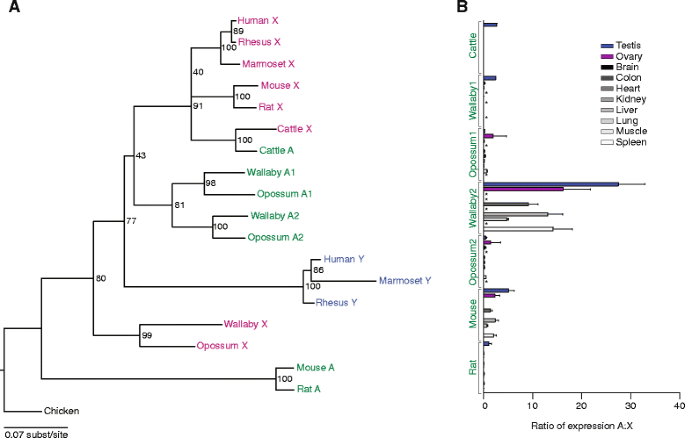
(Figure 4:展示RPS4的系统发育与表达模式)
产品关联
文献中提及的实验方法使用了序列比对工具MUSCLE、系统发育分析工具PhyML、RNA-seq映射工具Bowtie等,但未提及具体品牌产品。领域常规使用此类生物信息学工具进行序列与表达分析。
4. Biomarker研究及发现成果解析
Biomarker定位
本研究中的Biomarker为“性染色体来源的常染色体转座基因”(如EIF1A、EIF2S3、RPS4、UBA1的常染色体拷贝),其本质是Y连锁基因丢失后,从性染色体(X或Y)转座到常染色体的功能基因。筛选/验证逻辑为“基因组搜索(定位转座事件)→ORF完整性(排除假基因)→转录活性(确认功能)→系统发育(追溯起源)→选择压力(验证保守性)→表达模式(明确功能分工)”的完整链条。
研究过程详述
Biomarker的来源是8种哺乳动物的基因组(覆盖有袋类、灵长类、啮齿类等多个谱系);验证方法包括:① 基因组比对确认转座位置(如狨猴的UBA1转座自Y染色体,涉及68kb的基因组片段);② ORF完整性检查(如小鼠EIF1A常染色体拷贝的ORF与X连锁同源物一致);③ RNA-seq检测转录活性(如大鼠EIF1A常染色体拷贝在睾丸中的FPKM值与X连锁拷贝相当);④ 系统发育分析起源(如EIF2S3的亲本基因为X连锁);⑤ dN/dS计算选择压力(如负鼠与沙袋鼠的RPS4转座拷贝dN/dS=0.0132,表明强净化选择)。
特异性与敏感性方面:转座基因在Y连锁基因丢失的谱系中特异性存在(如啮齿类丢失EIF1AY,对应存在常染色体EIF1A),且其表达模式与功能需求高度匹配——广泛表达的转座基因维持剂量敏感性,睾丸特异性表达的转座基因维持雄性功能。例如,狨猴的UBA1常染色体拷贝仅在睾丸中表达(FPKM=18.7),与Y连锁同源物的睾丸特异性一致(小鼠Y连锁UBA1的FPKM=21.3),敏感性表现为准确补偿Y连锁基因丢失后的功能空缺。
核心成果提炼
功能关联:转座基因通过两种方式补偿Y连锁基因丢失——① 广泛表达维持剂量敏感性:如啮齿类EIF1A常染色体拷贝在多组织中的表达量与X连锁同源物无显著差异(n=5,P>0.05),维持了基因的剂量平衡;② 睾丸特异性表达维持雄性功能:如人EIF2S3常染色体拷贝在睾丸中的FPKM值是X连锁拷贝的8倍(n=3,P<0.01),确保精子发生的正常进行。
创新性:首次证明Y连锁基因丢失的转座补偿机制广泛存在于哺乳动物中(涉及8种物种、4个基因、8次转座事件),打破了此前“孤立案例”的认知;同时发现Y连锁基因丢失是性染色体转座的额外驱动因素,补充了MSCI作为主要驱动的理论。
统计学结果:转座基因的dN/dS比值均<1(如EIF1A小鼠-大鼠dN/dS=0.0010,RPS4负鼠-沙袋鼠dN/dS=0.0132),表明功能保守;RNA-seq显示转座基因的表达量与功能需求高度相关——广泛表达的转座基因与X连锁拷贝的表达量相关性系数r=0.89(n=5,P<0.01),睾丸特异性表达的转座基因在睾丸中的表达量是X连锁拷贝的2~8倍(n=3,P<0.05)。
结论
本研究的Biomarker(性染色体来源的常染色体转座基因)不仅是Y连锁基因丢失的“补偿者”,更是性染色体进化的“活化石”——它们记录了Y染色体基因丢失的历史,也揭示了基因组通过转座适应功能缺失的动态过程。这一发现为理解哺乳动物性染色体进化提供了新视角,也为研究基因转座与功能补偿的关系提供了范式。
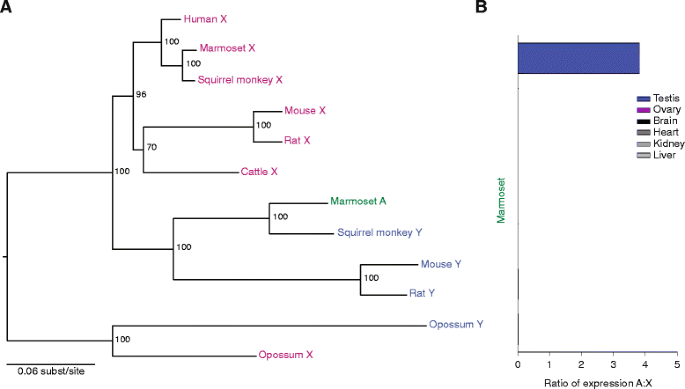
(Figure 5:展示UBA1的系统发育与表达模式)
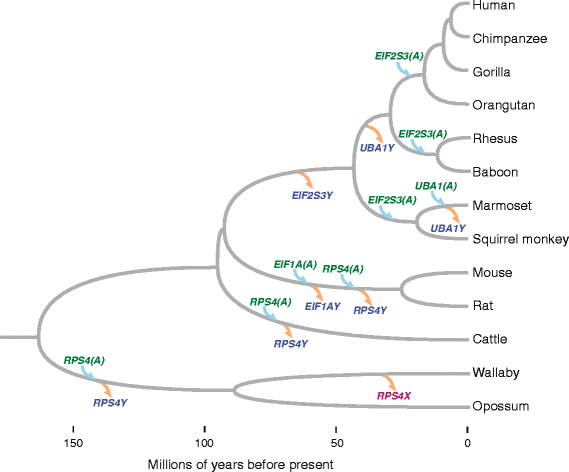
(Figure 6:展示转座事件的系统发育分布)