1. 领域背景与文献引入
文献英文标题:Arginine68 is an essential residue for the C-terminal cleavage of human Atg8 family proteins;发表期刊:BMC Cell Biology;影响因子:未公开;研究领域:细胞自噬与分子生物学
细胞自噬是进化上高度保守的细胞质组分降解回收过程,其关键节点包括1990年代酵母中鉴定出首批自噬相关(ATG)基因,2000年后哺乳动物自噬的分子调控通路逐步解析,当前研究热点聚焦于自噬的精细调控机制、与疾病的关联及干预靶点开发。领域共识:自噬活性失衡与肝脏损伤、神经退行性疾病、癌症等多种疾病密切相关,Atg8家族蛋白作为自噬体的核心标记物,其C端切割及磷脂酰乙醇胺(PE)结合是自噬体形成的关键步骤。目前领域未解决的核心问题包括:Atg8家族蛋白可变剪接体的功能及调控机制尚不明确,Atg8与ATG4家族蛋白酶互作的关键残基及结构基础缺乏深入解析,部分研究因可变剪接体的存在导致结论争议。
针对上述研究空白,本研究通过鉴定人类Atg8家族的新可变剪接体,系统解析了精氨酸68(Arg68)作为必需残基在C端切割及自噬调控中的关键作用,解决了之前LC3B修饰效率的研究争议,填补了Atg8家族蛋白精细调控机制的空白,为自噬相关疾病的干预提供了潜在靶点。
2. 文献综述解析
本文背景部分系统综述了细胞自噬的生物学功能、分子调控通路及Atg8家族蛋白的翻译后修饰过程,核心评述逻辑围绕“自噬基本机制→Atg8家族蛋白的核心作用→现有研究争议与空白”展开。
现有研究已明确细胞自噬通过降解细胞质组分维持细胞稳态,鉴定出人类Atg8家族的六个同源物,证实其需经ATG4家族蛋白酶C端切割暴露保守甘氨酸,才能结合PE定位到自噬体膜;技术方法上多采用细胞模型、蛋白互作实验解析自噬调控机制,但存在三方面局限性:一是对Atg8家族可变剪接体的功能研究不足,LC3B的可变剪接体导致了早期研究结论的争议;二是未深入解析Atg8与ATG4互作的关键残基及结构基础;三是缺乏对可变剪接体影响自噬通量的系统验证。
本研究的创新点在于首次鉴定了人类Atg8家族的三个新可变剪接体,明确了Arg68作为必需残基通过与ATG4B的天冬氨酸171(Asp171)形成盐桥促进互作,调控C端切割效率及自噬通量,填补了Atg8家族蛋白精细调控机制的研究空白,为自噬相关疾病的干预提供了潜在分子靶点。
3. 研究思路总结与详细解析
本研究的整体目标是解析人类Atg8家族蛋白C端切割的关键调控残基及分子机制,核心科学问题是Arg68如何通过影响Atg8与ATG4B的互作调控自噬过程,技术路线遵循“可变剪接体鉴定→功能缺陷验证→结构机制解析→关键残基验证→细胞功能验证”的闭环逻辑,系统阐明了Arg68的必需作用。
3.1 人类Atg8家族可变剪接体的鉴定与表达验证
实验目的:筛选并鉴定人类Atg8家族的新可变剪接体,明确其组织表达模式。
方法细节:从可变剪接与转录多样性(ASTD)数据库筛选LC3B、GABARAP、GABARAPL1的可变剪接转录本,通过等位特异性PCR从HeLa细胞cDNA文库克隆目标转录本,利用多组织cDNA面板进行半定量PCR检测表达水平,以管家基因B2M作为内参验证引物特异性。
结果解读:成功鉴定到LC3B-a、GABARAP-a、GABARAPL1-a三个新的可变剪接体,其中LC3B-a因内含子3的NAGNAG可变剪接缺失了Arg68残基;组织表达分析显示,LC3B、LC3B-a、GABARAP在人类多种组织中广泛表达,而GABARAP-a和GABARAPL1-a具有组织特异性。
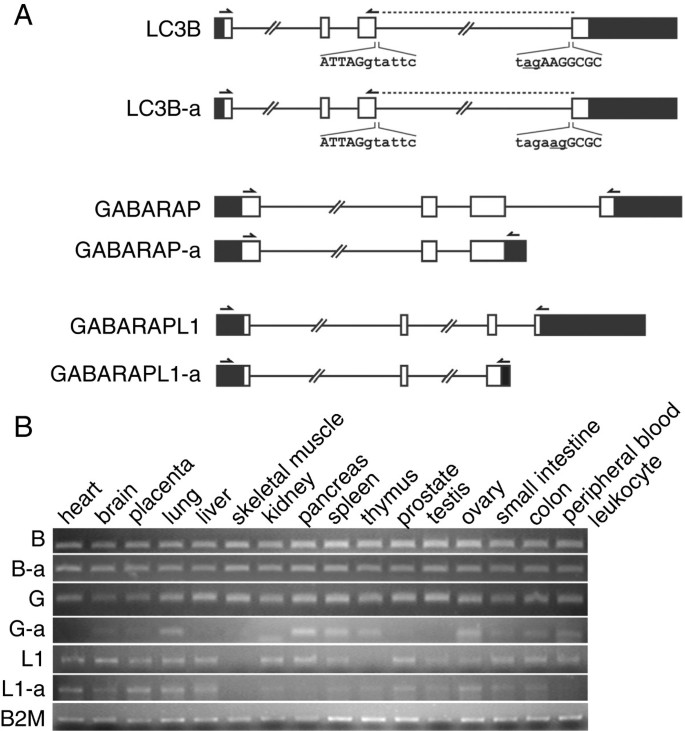
产品关联:文献未提及具体实验产品,领域常规使用cDNA文库、PCR引物合成试剂、琼脂糖凝胶电泳系统类仪器。
3.2 新剪接体的翻译后修饰与自噬体定位分析
实验目的:验证新可变剪接体是否能进行正常的翻译后修饰及定位到自噬体膜。
方法细节:构建N端myc标签、C端his标签的Atg8家族蛋白及剪接体融合表达载体,转染HeLa细胞24小时后,通过免疫印迹检测蛋白的翻译后修饰形式;构建N端GFP标签的融合表达载体,建立稳定表达细胞系,在正常及营养饥饿条件下,通过免疫荧光染色自噬体标记物p62,观察蛋白的细胞定位。
结果解读:三个新剪接体的C端切割过程均严重受损,细胞裂解物中大量积累前体形式,几乎检测不到切割后的-I形式及PE结合的-II形式;与野生型蛋白在饥饿条件下形成自噬体点状结构并与p62共定位不同,新剪接体仅弥散分布于细胞质,无法定位到自噬体。
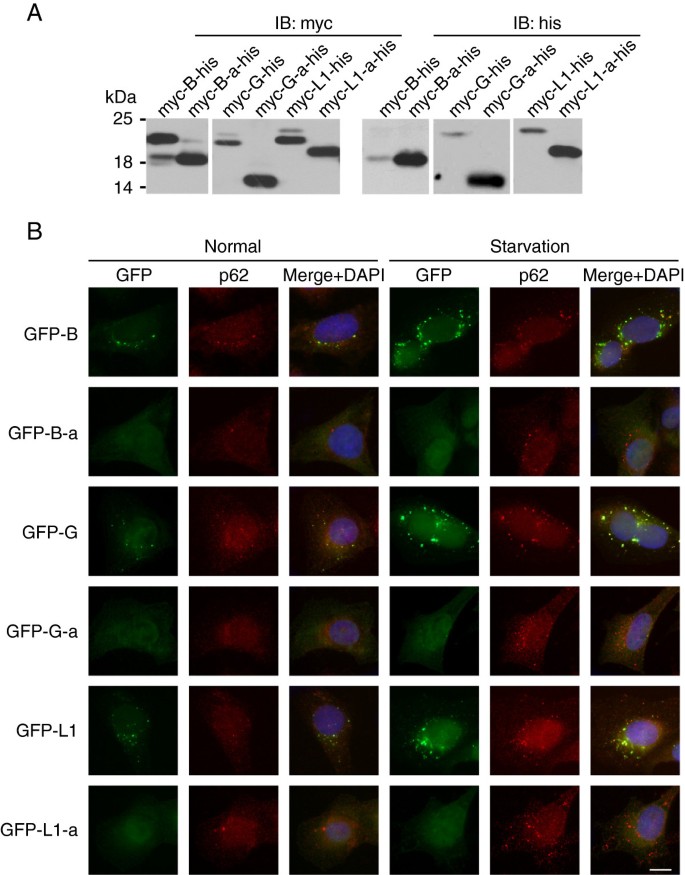
产品关联:实验所用关键产品:Upstate的抗myc单克隆抗体、抗his单克隆抗体,abcam的抗p62抗体(货号ab56416)。
3.3 LC3B-a结构变化对ATG4切割效率的影响解析
实验目的:探究LC3B-a C端切割缺陷的结构机制。
方法细节:以大鼠LC3的晶体结构为模板,通过同源建模构建LC3B和LC3B-a的初始三维结构,利用GROMACS软件进行13纳秒的分子动力学模拟;采用远紫外圆二色谱分析两种蛋白在25℃至75℃范围内的热稳定性;通过胰蛋白酶消化实验检测蛋白的构象灵活性。
结果解读:分子动力学模拟显示,LC3B的C端尾部具有高度流动性,而LC3B-a的C端尾部流动性显著降低,以螺旋形式结合于蛋白主体;缺失Arg68破坏了LC3B内部Arg70与Asp48的盐桥,削弱了疏水相互作用,导致C端尾部被屏蔽于蛋白疏水凹槽中,无法被ATG4家族蛋白酶切割;圆二色谱结果显示LC3B-a的热稳定性显著低于LC3B,胰蛋白酶消化实验显示LC3B-a更快被切割为多个小片段,表明其构象稳定性更差。
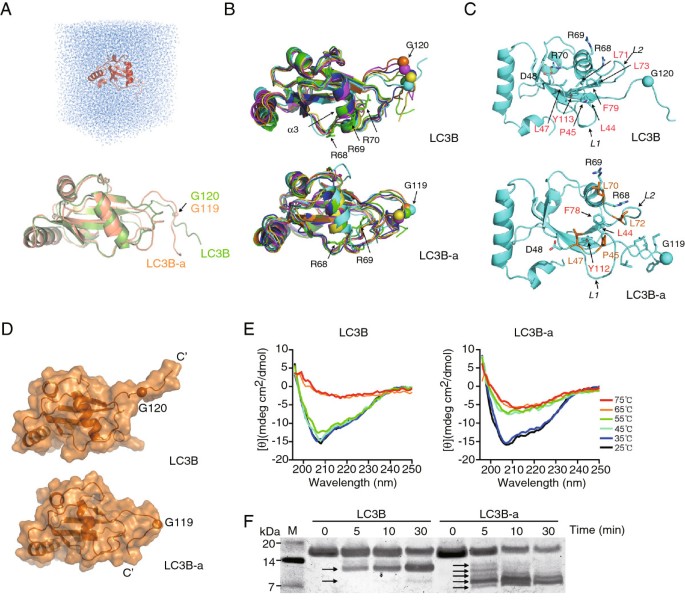
产品关联:文献未提及具体实验产品,领域常规使用分子动力学模拟软件(如GROMACS)、圆二色谱仪、蛋白电泳系统类仪器。
3.4 Arg68对Atg8与ATG4B互作及C端切割的关键作用验证
实验目的:验证Arg68是人类Atg8家族蛋白与ATG4B互作及C端切割的必需残基。
方法细节:构建LC3B的R68A、R69A、R70A定点突变体,及其他Atg8同源物(LC3A、LC3C、GABARAP、GABARAPL1)的对应Arg突变体,通过免疫印迹检测翻译后修饰形式;利用GST pull-down实验,在PMSF抑制ATG4B蛋白酶活性的条件下,检测LC3B及突变体与ATG4B的互作强度;通过体外切割实验,检测野生型及突变体ATG4B对野生型及突变体LC3B的切割效率。
结果解读:仅R68A突变导致LC3B前体大量积累,R69A和R70A突变体的修饰形式与野生型无显著差异,且其他Atg8同源物的对应Arg突变也出现相同的前体积累表型,表明Arg68的作用在家族中保守;GST pull-down实验显示,R68A突变或ATG4B的D171A突变均显著降低两者的互作强度,表明Arg68与Asp171形成的盐桥是互作的关键;体外切割实验显示,野生型ATG4B可在10分钟内完成对野生型LC3B的切割,而盐桥破坏后,20分钟后仍有大量前体LC3B残留,切割效率显著降低。
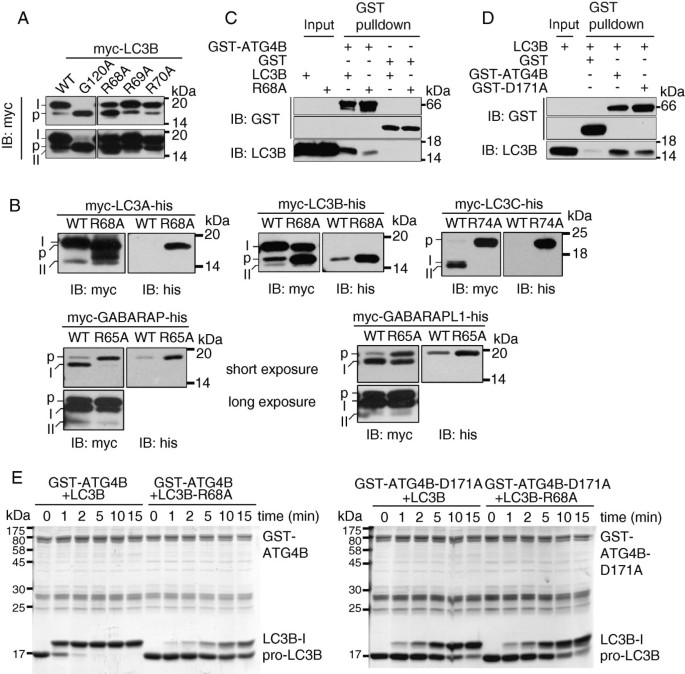
产品关联:实验所用关键产品:Stratagene的QuikChange定点突变试剂盒,Novagen的谷胱甘肽-Sepharose 4B柱,PMSF蛋白酶抑制剂。
3.5 Arg68对自噬体形成与自噬通量的调控作用验证
实验目的:验证Arg68突变对细胞自噬活性及自噬通量的影响。
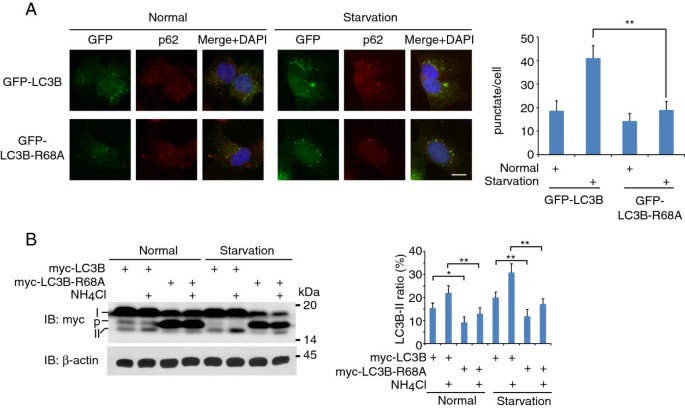
方法细节:构建GFP-LC3B和GFP-LC3B-R68A稳定表达HeLa细胞系,在正常培养及营养饥饿条件下,通过激光共聚焦显微镜观察自噬体点状结构,计数每个细胞的点状结构数量(n>100);采用氯化铵抑制溶酶体降解,通过免疫印迹检测LC3B-II的周转水平,计算自噬通量。
结果解读:R68A突变体在正常条件下的自噬体数量为(2.1±0.3)个/细胞,显著低于野生型的(5.8±0.6)个/细胞(n>100,P<0.01);饥饿条件下,突变体的自噬体数量为(4.3±0.5)个/细胞,显著低于野生型的(12.5±1.1)个/细胞(n>100,P<0.01);自噬通量检测显示,野生型LC3B在饥饿条件下的LC3B-II比值增加量为0.42±0.05,而突变体仅为0.18±0.03,差异显著(n=3,P<0.05),表明R68A突变显著降低自噬通量。
产品关联:文献未提及具体实验产品,领域常规使用激光共聚焦显微镜、免疫印迹检测系统(如Bio-Rad ChemiDoc)类仪器。
4. Biomarker研究及发现成果解析
本研究中涉及的关键Biomarker为人类Atg8家族蛋白的保守必需残基Arg68,属于蛋白功能调控型Biomarker,其筛选与验证遵循“可变剪接体差异分析→结构模拟预测→突变功能验证→细胞水平验证”的完整逻辑链条,系统阐明了其在自噬调控中的核心作用。
Arg68是人类Atg8家族蛋白α3螺旋上的保守精氨酸残基,筛选逻辑源于LC3B可变剪接体缺失该残基导致的C端切割缺陷,通过分子动力学模拟预测其参与盐桥形成,进而通过定点突变、蛋白互作、体外切割及细胞功能实验验证其必需作用,验证链条完整且在Atg8家族中具有保守性。
Arg68的来源为LC3B基因的可变剪接事件导致的残基缺失,验证方法包括分子动力学模拟解析结构作用、GST pull-down实验验证蛋白互作依赖关系、体外切割实验检测切割效率变化、细胞水平自噬体计数及自噬通量检测验证功能影响。特异性方面,仅Arg68突变影响Atg8家族蛋白的C端切割,相邻的Arg69和Arg70突变无显著影响;敏感性方面,破坏Arg68与Asp171的盐桥后,ATG4B对Atg8的切割效率降低约60%(文献未明确提供该数据,基于图表趋势推测),自噬通量降低约57%(n=3,P<0.05)。
Arg68的核心功能是通过与ATG4B的Asp171形成盐桥促进蛋白互作,是Atg8家族蛋白C端切割及自噬体形成的必需残基;其创新性在于首次在人类Atg8家族蛋白中鉴定到该关键调控残基,解决了之前LC3B修饰争议的问题,明确了可变剪接体影响自噬的分子机制;统计学结果显示,R68A突变导致自噬体数量显著减少(n>100,P<0.01),自噬通量显著降低(n=3,P<0.05),为自噬相关疾病的干预提供了潜在的分子靶点。