1. 领域背景与文献
文献英文标题:Zinc finger protein ZNF33B facilitates Japanese encephalitis virus replication by regulating m⁶A modification on Ifih1 and Irf3 transcripts to antagonize antiviral response;发表期刊:Cell & Bioscience;影响因子:未公开;研究领域:病毒免疫学(黄病毒感染与天然免疫调控)。
日本脑炎病毒是黄病毒科黄病毒属的嗜神经RNA病毒,主要通过库蚊叮咬传播,流行于亚洲及西太平洋地区,全球每年约报告6.8万临床病例,病死率为10%~30%,30%~50%的幸存者会遗留帕金森病、癫痫、认知障碍等永久性神经后遗症,对公共卫生安全造成严重威胁。领域共识:RIG-I样受体介导的天然免疫通路是宿主抗RNA病毒感染的第一道防线,病毒RNA被RIG-I样受体识别后,通过线粒体抗病毒信号蛋白招募下游激酶,激活干扰素调节因子3,进而诱导I型干扰素及干扰素刺激基因的表达,启动抗病毒免疫应答。现有研究已证实黄病毒可通过蛋白酶切割、泛素化降解等方式靶向RIG-I样受体通路关键分子实现免疫逃逸,但宿主因子介导的表观转录组层面调控机制尚未被充分阐明。
锌指蛋白是人类基因组中最大的转录因子家族之一,传统认为其主要通过结合DNA调控基因转录,近年研究发现部分锌指蛋白可结合RNA,参与RNA剪接、出核、稳定性调控等多种转录后调控过程。锌指蛋白33B(ZNF33B)已被报道参与胚胎发育、器官发生、细胞分化等生理过程,且在神经组织中高表达,但其在嗜神经病毒感染中的功能及机制仍不明确。N6-甲基腺苷(m⁶A)是真核mRNA中最丰富的内部化学修饰,由甲基转移酶复合物(Writer)、去甲基化酶(Eraser)和甲基化阅读蛋白(Reader)动态调控,参与RNA代谢的多个环节。已有研究显示寨卡病毒、丙肝病毒等黄病毒可劫持宿主m⁶A通路调控病毒复制或逃逸免疫识别,但日本脑炎病毒感染中m⁶A修饰的作用,以及锌指蛋白是否可作为衔接分子协调m⁶A通路与天然免疫调控,仍有待阐明。本研究针对上述研究空白,旨在揭示ZNF33B调控日本脑炎病毒复制的分子机制,明确其对RIG-I样受体通路和m⁶A修饰的调控作用,为抗黄病毒药物研发提供新的靶点。
2. 文献综述解析
作者的文献综述按照“病毒-宿主免疫互作→锌指蛋白功能扩展→m⁶A修饰的免疫调控作用”三个维度进行分类梳理,系统总结了领域内现有研究的进展与局限性,明确本研究的创新定位。
在黄病毒与宿主天然免疫互作研究领域,现有研究已证实RIG-I样受体通路是抑制黄病毒复制的核心信号轴,黄病毒可通过编码非结构蛋白直接靶向RIG-I、线粒体抗病毒信号蛋白等分子,通过蛋白酶水解、泛素化降解等方式阻断信号传导,该类研究的优势是明确了病毒蛋白介导的直接免疫逃逸机制,但局限性在于较少关注宿主内源性因子对免疫通路的转录后调控,尤其是表观转录组层面的调控网络。在锌指蛋白功能研究领域,现有研究已发现C2H2型锌指蛋白不仅具有DNA结合活性,还可通过其锌指结构结合RNA,参与核糖体质量控制、RNA降解等多种转录后调控过程,ZNF33B已被证实参与哮喘易感性、脓毒症预后等病理过程,且该分子在大脑皮层、海马等神经组织中高表达,提示其可能参与嗜神经病毒的感染调控,但现有研究尚未明确其在抗病毒天然免疫中的具体功能。在m⁶A修饰的免疫调控研究领域,现有研究发现m⁶A修饰可通过调控抗病毒基因的RNA代谢参与天然免疫应答,如丙肝病毒利用m⁶A阅读蛋白YTHDC2增强病毒内部核糖体进入位点介导的翻译,寨卡病毒通过调控宿主RNA的m⁶A修饰逃逸免疫识别,该类研究揭示了m⁶A通路在病毒感染中的重要作用,但尚未阐明宿主RNA结合蛋白是否可作为衔接分子,协调m⁶A修饰的沉积、出核及降解全过程,实现对天然免疫通路的系统性调控。
通过对比现有研究的局限性,本研究的创新价值主要体现在三个方面:首次发现ZNF33B是日本脑炎病毒免疫逃逸的关键宿主因子;首次揭示了锌指蛋白可作为衔接分子,同时协调m⁶A通路的Writer、核Reader和质Reader蛋白,实现对靶标RNA的多步骤调控;首次阐明了同时靶向RIG-I样受体通路多个关键分子转录本的免疫逃逸机制,为理解黄病毒与宿主互作提供了新的视角。
3. 研究思路总结与详细解析
本研究的核心目标是阐明ZNF33B调控日本脑炎病毒复制的分子机制,核心科学问题是ZNF33B如何通过调控抗病毒转录本的m⁶A修饰抑制RIG-I样受体介导的天然免疫应答,技术路线遵循“功能验证→分子互作→机制解析→通路验证”的闭环逻辑:首先明确ZNF33B对RIG-I样受体通路的负调控功能,随后验证其与抗病毒转录本、m⁶A通路蛋白的互作关系,进而阐明其调控m⁶A修饰、转录本出核及降解的具体分子机制,最终确认该调控轴对日本脑炎病毒复制的促进作用。
3.1 ZNF33B对RIG-I样受体天然免疫通路的负调控功能验证
实验目的:明确ZNF33B是否通过抑制RIG-I样受体通路促进日本脑炎病毒复制。
方法细节:采用人胚肾HEK293T细胞模型,分别过表达或敲除ZNF33B,结合日本脑炎病毒感染处理,通过RNA免疫沉淀、免疫印迹、实时荧光定量PCR、双荧光素酶报告实验等方法,检测RIG-I样受体通路关键分子的表达水平、干扰素启动子活性及病毒复制水平。
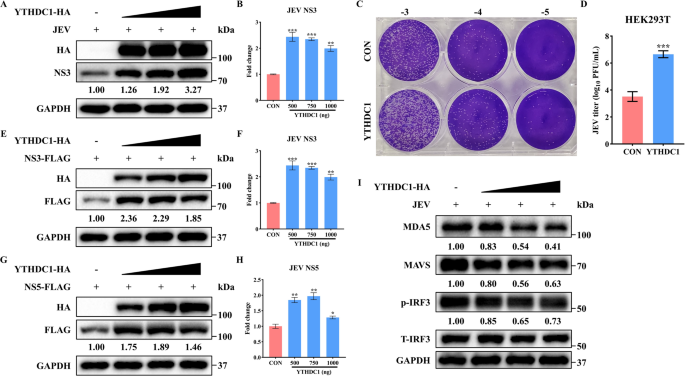
结果解读:首先通过RNA免疫沉淀和免疫荧光实验证实,日本脑炎病毒的RNA可被RIG-I和黑色素瘤分化相关蛋白5(MDA5)识别,富集倍数分别为对照组的4.2倍和3.8倍(n=3,P<0.001),验证日本脑炎病毒感染可有效激活RIG-I样受体通路。过表达ZNF33B后,日本脑炎病毒非结构蛋白NS3的蛋白水平上调2.3倍(n=3,P<0.01),提示病毒复制增强;同时MDA5、线粒体抗病毒信号蛋白、磷酸化干扰素调节因子3、磷酸化信号转导及转录激活蛋白1的蛋白水平显著下调,干扰素α2、干扰素β1、干扰素刺激基因15、干扰素刺激基因56的mRNA水平降低40%~60%(n=3,P<0.01),干扰素β启动子活性被抑制65%(n=3,P<0.001);敲除ZNF33B则得到相反结果,提示ZNF33B负调控RIG-I样受体介导的天然免疫应答。对应实验结果见图1。
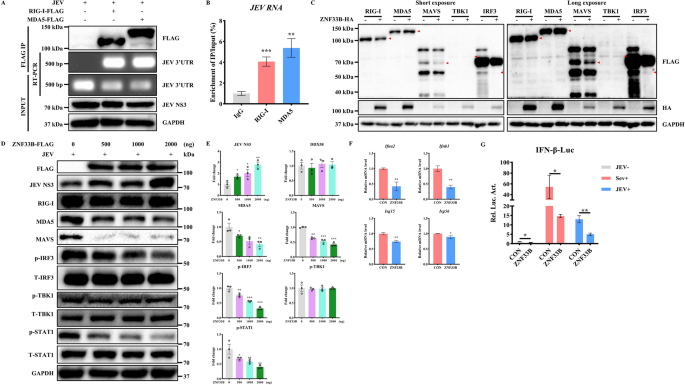
产品关联:实验所用关键产品:Proteintech的MDA5抗体(货号21775-1-AP)、Santa Cruz的线粒体抗病毒信号蛋白抗体(货号sc-166583)、Cell Signaling Technology的磷酸化干扰素调节因子3抗体(货号5483),Promega的双荧光素酶报告检测试剂盒。
3.2 ZNF33B与抗病毒转录本的结合及出核调控验证
实验目的:明确ZNF33B是否直接结合RIG-I样受体通路关键分子的转录本,并调控其核质分布与稳定性。
方法细节:在日本脑炎病毒感染的HEK293T细胞中过表达Flag标记的ZNF33B,通过RNA免疫沉淀结合实时荧光定量PCR检测ZNF33B结合的mRNA种类;采用核质分离实验检测抗病毒转录本在细胞核和细胞质中的分布差异;通过放线菌素D处理抑制细胞转录,在不同时间点检测转录本的表达水平,计算其半衰期。
结果解读:RNA免疫沉淀实验显示,ZNF33B可显著富集Ifih1(编码MDA5)、Mavs(编码线粒体抗病毒信号蛋白)、Irf3(编码干扰素调节因子3)的转录本,富集倍数分别为对照组的5.1倍、3.7倍和4.3倍(n=3,P<0.001)。核质分离实验显示,过表达ZNF33B后,细胞核内Ifih1、Mavs、Irf3的转录本水平降低40%~50%,细胞质内水平升高60%~80%(n=3,P<0.01),提示ZNF33B促进这些转录本的核输出。放线菌素D处理实验显示,ZNF33B过表达可显著缩短Ifih1和Irf3转录本的半衰期,Ifih1的半衰期从15.20h降至4.68h,Irf3的半衰期从13.43h降至5.63h(n=3,P<0.001);敲除ZNF33B则显著延长二者的半衰期,而Mavs转录本的稳定性无显著变化,提示ZNF33B特异性促进Ifih1和Irf3转录本的降解。对应实验结果见图2。
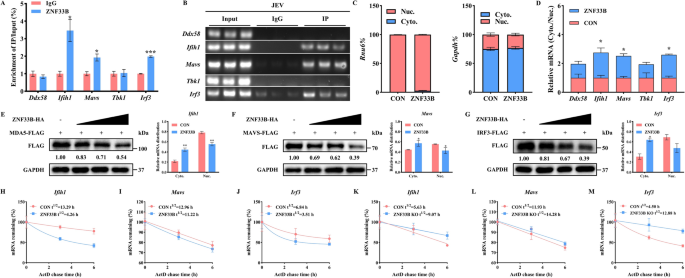
产品关联:实验所用关键产品:Norgen Biotek的核质RNA提取试剂盒,MCE的放线菌素D。
3.3 ZNF33B对抗病毒转录本m⁶A修饰的调控机制验证
实验目的:明确ZNF33B是否通过招募m⁶A甲基转移酶增强抗病毒转录本的m⁶A修饰水平。
方法细节:通过m⁶A甲基化RNA免疫沉淀结合实时荧光定量PCR(MeRIP-qPCR),检测日本脑炎病毒感染后抗病毒转录本的m⁶A修饰水平变化;通过免疫共沉淀实验检测ZNF33B与m⁶A甲基转移酶(甲基转移酶样3(METTL3)、甲基转移酶样14(METTL14))的相互作用;在METTL14敲低的细胞中,验证ZNF33B对RIG-I样受体通路分子的调控作用是否依赖METTL14。
结果解读:日本脑炎病毒感染可显著上调细胞总m⁶A水平,且Ifih1、Mavs、Irf3转录本的m⁶A修饰水平分别升高2.7倍、2.1倍和2.3倍(n=3,P<0.01)。免疫共沉淀实验证实,ZNF33B可直接结合METTL14,而不与METTL3互作。过表达ZNF33B可增强METTL3对Ifih1、Mavs、Irf3转录本的富集,富集倍数分别提升2.2倍、1.8倍和2.0倍(n=3,P<0.01)。敲低METTL14后,ZNF33B对MDA5、磷酸化干扰素调节因子3的下调作用被完全逆转,提示ZNF33B对RIG-I样受体通路的抑制依赖METTL14介导的m⁶A修饰。对应实验结果见图3。
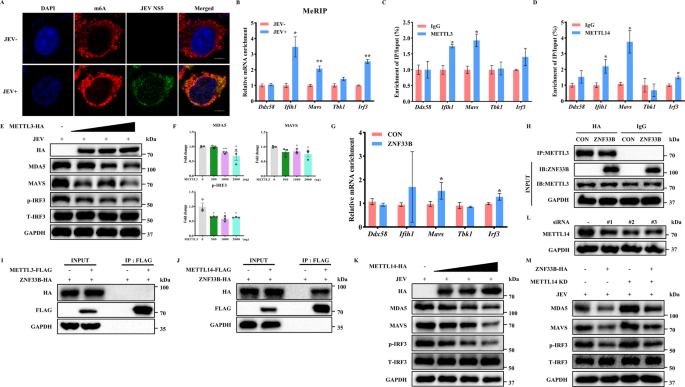
产品关联:实验所用关键产品:Proteintech的m⁶A抗体(货号68055-1-Ig)、METTL14抗体(货号26158-1-AP)。
3.4 m⁶A修饰后转录本的出核调控机制验证
实验目的:明确ZNF33B是否通过招募核m⁶A阅读蛋白YTH结构域包含蛋白1(YTHDC1),促进甲基化转录本的核输出。
方法细节:通过RNA免疫沉淀结合实时荧光定量PCR,检测YTHDC1结合的抗病毒转录本;通过免疫共沉淀和免疫荧光实验,验证ZNF33B与YTHDC1的相互作用与亚细胞共定位;在YTHDC1敲低的细胞中,检测抗病毒转录本的核质分布变化,验证YTHDC1在ZNF33B介导的转录本出核中的作用;通过空斑实验检测YTHDC1对日本脑炎病毒滴度的影响。
结果解读:RNA免疫沉淀实验显示,YTHDC1可结合Ifih1、Mavs、Irf3的转录本,富集倍数分别为对照组的3.9倍、2.8倍和3.5倍(n=3,P<0.001)。免疫共沉淀实验证实ZNF33B与YTHDC1存在直接相互作用,免疫荧光显示二者在细胞核内形成共定位的凝聚体。过表达YTHDC1可促进抗病毒转录本的核输出,使细胞质内Ifih1、Mavs、Irf3的转录本水平升高50%~70%(n=3,P<0.01);同时YTHDC1过表达可使日本脑炎病毒NS3蛋白水平上调1.9倍,病毒滴度升高2.2倍(n=3,P<0.01)。敲低YTHDC1后,ZNF33B诱导的抗病毒转录本核输出现象被逆转,核内Ifih1、Mavs、Irf3转录本水平恢复至对照组的80%以上,提示ZNF33B介导的转录本出核依赖YTHDC1。对应实验结果见图4和图5。
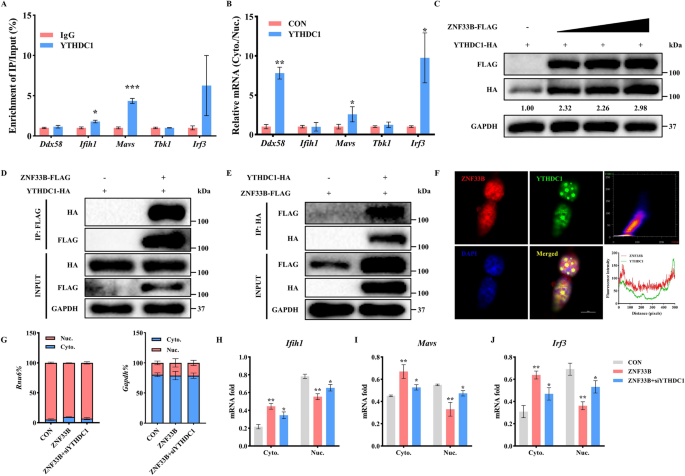
产品关联:实验所用关键产品:Proteintech的YTHDC1抗体(货号14392-1-AP),Invitrogen的Alexa Fluor系列荧光二抗。
3.5 细胞质中转录本的降解机制验证
实验目的:明确细胞质中m⁶A修饰的抗病毒转录本是否被YTH结构域家族蛋白2(YTHDF2)识别并降解,以及该过程是否依赖ZNF33B。
方法细节:通过RNA免疫沉淀结合实时荧光定量PCR,检测YTHDF2结合的抗病毒转录本;通过放线菌素D处理,检测YTHDF2对转录本半衰期的影响;在YTHDF2敲低的细胞中,验证ZNF33B对转录本稳定性的调控作用是否依赖YTHDF2;通过空斑实验检测YTHDF2对日本脑炎病毒滴度的影响。
结果解读:RNA免疫沉淀实验显示,YTHDF2可结合Ifih1和Irf3的转录本,富集倍数分别为对照组的4.5倍和3.7倍(n=3,P<0.001),对Mavs转录本无显著富集。过表达YTHDF2可显著缩短Ifih1和Irf3转录本的半衰期,分别从14.8h降至5.2h、13.1h降至6.1h(n=3,P<0.01),并下调MDA5和干扰素调节因子3的蛋白水平。免疫共沉淀实验证实ZNF33B可与YTHDF2相互作用,且过表达ZNF33B可增强YTHDF2对Ifih1和Irf3转录本的结合,富集倍数分别提升2.3倍和2.0倍(n=3,P<0.001)。敲低YTHDF2后,ZNF33B诱导的Ifih1和Irf3转录本降解被逆转,其半衰期分别从4.68h恢复至7.92h、5.63h恢复至8.47h(n=3,P<0.01)。过表达YTHDF2可显著上调日本脑炎病毒NS3和NS5的蛋白水平,使病毒滴度升高1.8倍(n=3,P<0.01),促进病毒复制。对应实验结果见图6。
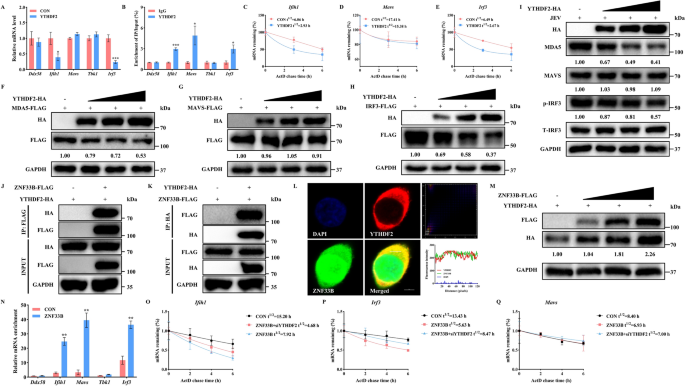
产品关联:实验所用关键产品:Proteintech的YTHDF2抗体(货号24744-1-AP),碧云天的Protein A+G磁珠。
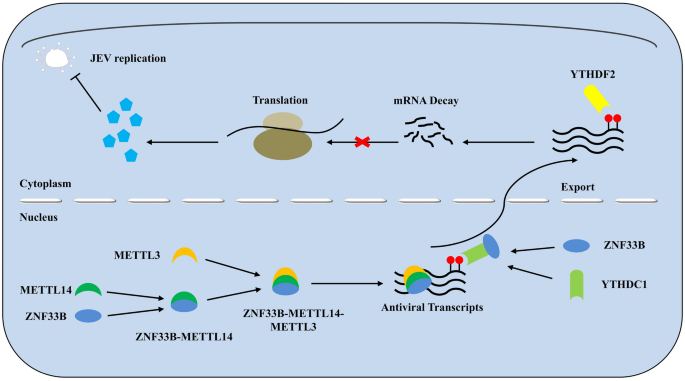
本研究最终提出的调控机制模型如图7所示,日本脑炎病毒感染后,ZNF33B在细胞核内招募METTL14,增强Ifih1和Irf3转录本的m⁶A修饰,随后结合YTHDC1促进甲基化转录本出核,进入细胞质后被YTHDF2识别并降解,最终抑制RIG-I样受体介导的抗病毒免疫应答,促进病毒复制。
4. Biomarker研究及发现成果
本研究未涉及临床生物标志物的筛选与验证,主要聚焦于病毒感染的分子机制解析,明确了ZNF33B-m⁶A调控轴可作为日本脑炎病毒感染的潜在干预靶点,其功能与调控特征如下。
靶点定位:本研究发现的核心功能性靶点为锌指蛋白33B(ZNF33B),其调控的效应分子为Ifih1和Irf3转录本的m⁶A修饰,调控逻辑链条完整:日本脑炎病毒感染后ZNF33B发挥作用,依次招募METTL14增加靶转录本的m⁶A修饰、结合YTHDC1促进转录本核输出、协同YTHDF2介导转录本降解,最终实现对RIG-I样受体通路的系统性抑制。
研究过程详述:ZNF33B的功能验证基于HEK293T、猪肾PK-15等细胞系的日本脑炎病毒感染模型,通过免疫印迹和实时荧光定量PCR检测其表达变化;m⁶A修饰的检测采用MeRIP-qPCR方法,在日本脑炎病毒感染细胞中,Ifih1转录本的m⁶A富集倍数为2.7倍(n=3,P<0.01),Irf3转录本的m⁶A富集倍数为2.3倍(n=3,P<0.01);功能验证显示,过表达ZNF33B可使日本脑炎病毒滴度升高2.1倍(n=3,P<0.001),敲低ZNF33B则使病毒滴度降低62%(n=3,P<0.01)。
核心成果提炼:本研究首次发现ZNF33B是日本脑炎病毒免疫逃逸的关键宿主因子,可作为衔接分子协调m⁶A通路的Writer、核Reader和质Reader蛋白,同时靶向RIG-I样受体通路多个关键分子的转录本,实现对天然免疫应答的全面抑制。该调控轴的干预可显著抑制日本脑炎病毒复制(P<0.001,n=3),为抗黄病毒药物研发提供了全新的靶点,同时拓展了锌指蛋白和m⁶A修饰在抗病毒免疫调控中的功能认知。