1. 领域背景与文献
文献英文标题:3D epigenome architecture orchestrates cis and trans regulation of flowering time in rice;发表期刊:Genome Biology;影响因子:未公开;研究领域:植物三维基因组学、水稻开花时间表观遗传调控。
领域共识:真核生物的线性染色质在细胞核内折叠形成复杂的三维结构,广泛参与基因转录、DNA复制、DNA损伤修复等核心生物学过程。该领域的关键发展节点包括:2009年高通量染色质构象捕获技术首次绘制人类基因组高分辨率三维互作图谱,明确了染色质环、拓扑相关结构域等高级结构单元;2019年水稻中首次绘制了组蛋白H3赖氨酸4三甲基化、组蛋白H3赖氨酸9二甲基化等修饰相关的染色质互作图谱,揭示了三维结构对基因转录的调控功能。当前研究热点主要集中于三维基因组架构与表观遗传修饰的协同调控机制,以及液-液相分离在染色质高级结构形成中的作用。目前领域内未解决的核心问题包括:水稻中抑制型表观标记组蛋白H3赖氨酸27三甲基化相关的三维基因组架构特征尚不明确,其如何调控开花等重要农艺性状的分子机制也未被解析。本研究针对上述研究空白,通过绘制全基因组组蛋白H3赖氨酸27三甲基化相关的染色质互作图谱,解析其对水稻开花时间的调控机制,为水稻开花性状的分子设计育种提供新的理论靶点。
2. 文献综述解析
作者对现有研究的评述逻辑按照研究主题分为三个维度,分别为三维基因组的功能研究、水稻开花调控分子网络、组蛋白H3赖氨酸27三甲基化的表观遗传调控作用。
现有研究中,三维基因组领域已证实动物和植物中染色质高级结构可介导基因的协同转录调控,例如果蝇胚胎发育过程中同源基因通过形成染色质环维持共表达,水稻中干旱响应基因通过组蛋白H3赖氨酸9乙酰化标记的启动子-启动子互作实现协同表达,此类研究的优势是明确了染色质互作的转录调控功能,但局限性在于多集中于活性组蛋白修饰相关的互作,对抑制型修饰组蛋白H3赖氨酸27三甲基化相关的三维互作研究较少。水稻开花调控领域已克隆超过110个开花相关基因,明确了“水稻 gigantea-抽穗期1号-抽穗期3a/水稻开花素T1”和“早抽穗1号-抽穗期3a/水稻开花素T1”两条核心调控通路,优势是阐明了单个基因的功能和线性调控关系,局限性是未从三维基因组层面解释多个开花关键基因的协同调控机制。组蛋白H3赖氨酸27三甲基化调控领域已证实该标记由多梳抑制复合物2催化,水稻中多梳抑制复合物2组分如水稻胚胎花2b、水稻类卷曲叶基因等可通过调控组蛋白H3赖氨酸27三甲基化水平抑制开花相关基因表达,优势是明确了该修饰的表观调控作用,局限性是未揭示其是否通过形成高级染色质结构实现多基因的协同抑制。
本研究的创新价值在于,首次在全基因组水平绘制了水稻组蛋白H3赖氨酸27三甲基化相关的三维染色质互作图谱,发现了开花关键基因早抽穗1号、抽穗期3a、水稻开花素T1形成的跨染色体抑制型空间簇,并揭示了液-液相分离介导的反式调控复合物对该簇的调控机制,填补了三维表观基因组调控水稻开花的研究空白,为理解植物中多基因协同转录调控提供了新的范式。
3. 研究思路总结与详细解析
本研究的核心目标是解析组蛋白H3赖氨酸27三甲基化相关的三维表观基因组架构对水稻开花时间的调控机制,核心科学问题包括:水稻中组蛋白H3赖氨酸27三甲基化相关的染色质高级结构具有哪些特征?该结构如何参与开花关键基因的协同调控?反式调控因子如何结合该结构实现开花时间的精准控制?技术路线遵循“结构发现-动态验证-功能解析-机制阐明”的闭环逻辑:首先通过长读长染色质相互作用配对末端标签测序技术绘制组蛋白H3赖氨酸27三甲基化相关的染色质互作图谱,鉴定其结构特征;其次通过多组织、多品种比较分析该结构的动态性与稳定性;接着通过突变体验证多梳抑制复合物2组分对该结构的调控作用;随后通过分子生物学实验验证开花调控复合物对关键基因簇的结合;最后通过液-液相分离实验解析反式调控的分子机制。
3.1 组蛋白H3赖氨酸27三甲基化相关染色质互作图谱的绘制与特征分析
实验目的:明确水稻中组蛋白H3赖氨酸27三甲基化相关的染色质互作的全基因组特征。
方法细节:采用长读长染色质相互作用配对末端标签测序技术,使用组蛋白H3赖氨酸27三甲基化抗体,分别对明恢63、珍汕97两个水稻品种的叶和根组织,以及东津野生型和水稻胚胎花2b突变体的叶组织进行检测,设置生物学重复,通过与已发表的组蛋白H3赖氨酸27三甲基化染色质免疫共沉淀测序数据比对验证数据质量。
结果解读:在明恢63叶、明恢63根、珍汕97叶、珍汕97根中分别鉴定到13746、15372、15181、15313个高置信度染色质互作(错误发现率<0.05),这些互作连接了40%-57%的组蛋白H3赖氨酸27三甲基化标记区域(称为锚点),锚点区域的组蛋白H3赖氨酸27三甲基化信号强度显著高于未参与互作的基础峰,且锚点基因的表达水平显著低于基础基因。通过免疫荧光实验观察到组蛋白H3赖氨酸27三甲基化在细胞核中形成大量斑点,部分与组蛋白H3赖氨酸4三甲基化的斑点重叠,对应结果如下图所示:
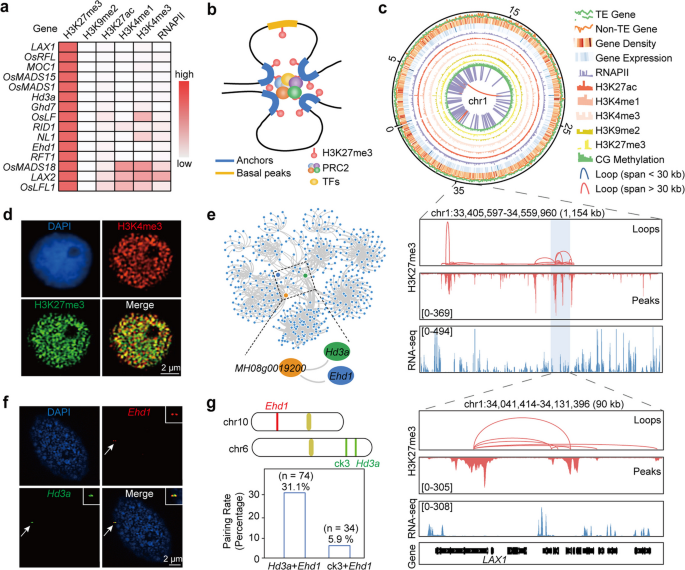
产品关联:实验所用关键产品:ABclonal的组蛋白H3赖氨酸27三甲基化抗体(货号A2363)。
3.2 抑制型染色质空间簇的鉴定与开花相关簇的验证
实验目的:解析组蛋白H3赖氨酸27三甲基化相关的染色质高级结构特征,鉴定与开花调控相关的空间簇。
方法细节:基于长读长染色质相互作用配对末端标签测序数据的连接性和接触频率,识别染色质空间簇;通过杂交链式反应荧光原位杂交技术,使用早抽穗1号和抽穗期3a的特异性探针,在明恢63叶细胞核中验证两个基因的空间共定位。
结果解读:共鉴定到342个组蛋白H3赖氨酸27三甲基化相关的抑制型染色质空间簇,其中发现关键开花基因早抽穗1号(位于10号染色体)和抽穗期3a/水稻开花素T1(位于6号染色体)形成跨染色体的空间簇,杂交链式反应荧光原位杂交实验显示早抽穗1号和抽穗期3a的荧光信号重叠率显著高于阴性对照(n=74个细胞核),证实二者的空间邻近,对应结果如3.1部分图1f、1g所示。
产品关联:文献未提及具体实验产品,领域常规使用荧光原位杂交类试剂与探针合成服务。
3.3 组蛋白H3赖氨酸27三甲基化染色质环的动态性分析
实验目的:明确组蛋白H3赖氨酸27三甲基化相关染色质互作在不同组织、不同品种中的动态变化特征。
方法细节:比较明恢63叶和根的长读长染色质相互作用配对末端标签测序数据,鉴定组织特异性的互作环和抑制型染色质空间簇;比较明恢63和珍汕97叶的测序数据,分析品种特异性的互作差异,结合基因组变异分析差异的遗传基础。
结果解读:叶中存在1254个特异性互作环,根中存在4070个特异性互作环,组织特异性环相关的锚点基因在对应组织中的表达水平显著更低;不同品种间的差异组蛋白H3赖氨酸27三甲基化峰中,30.5%存在存在/缺失变异,44.1%存在单核苷酸多态性,提示遗传变异是组蛋白H3赖氨酸27三甲基化互作差异的重要原因。同时发现早抽穗1号-抽穗期3a/水稻开花素T1空间簇在不同组织、不同品种中均稳定存在,仅连接的桥基因存在差异,且两个基因的组蛋白H3赖氨酸27三甲基化水平在四个组织(幼叶、根、穗、成熟叶)中呈显著正相关(明恢63中相关系数≥0.88,珍汕97中相关系数=0.82),对应结果如下图所示:
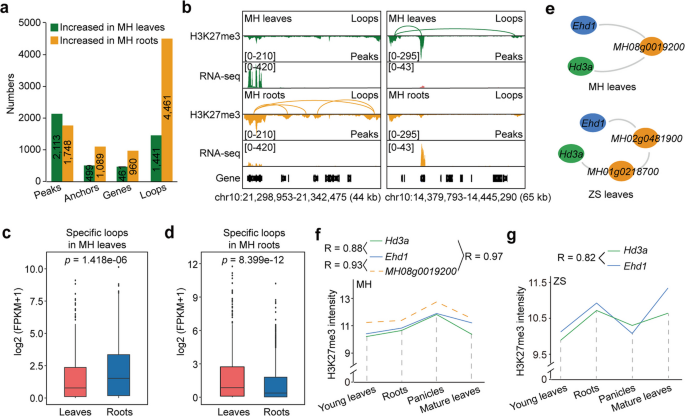
产品关联:文献未提及具体实验产品,领域常规使用高通量测序数据分析工具。
3.4 水稻胚胎花2b对组蛋白H3赖氨酸27三甲基化染色质互作的调控作用验证
实验目的:验证多梳抑制复合物2组分水稻胚胎花2b对组蛋白H3赖氨酸27三甲基化相关染色质互作的调控作用。
方法细节:构建水稻胚胎花2b的转移DNA插入突变体(东津背景),分别对野生型和突变体的叶组织进行组蛋白H3赖氨酸27三甲基化染色质免疫共沉淀测序和长读长染色质相互作用配对末端标签测序检测,比较二者的染色质互作差异;通过杂交链式反应荧光原位杂交验证突变体中早抽穗1号和抽穗期3a的空间共定位。
结果解读:水稻胚胎花2b突变体中组蛋白H3赖氨酸27三甲基化相关的染色质互作网络发生显著重塑,丢失了2905个组蛋白H3赖氨酸27三甲基化锚点,新增834个锚点,但早抽穗1号-抽穗期3a/水稻开花素T1空间簇仍然存在,仅整合了更多的桥基因,杂交链式反应荧光原位杂交实验证实突变体中二者仍存在空间共定位(n=42个细胞核),对应结果如下图所示:
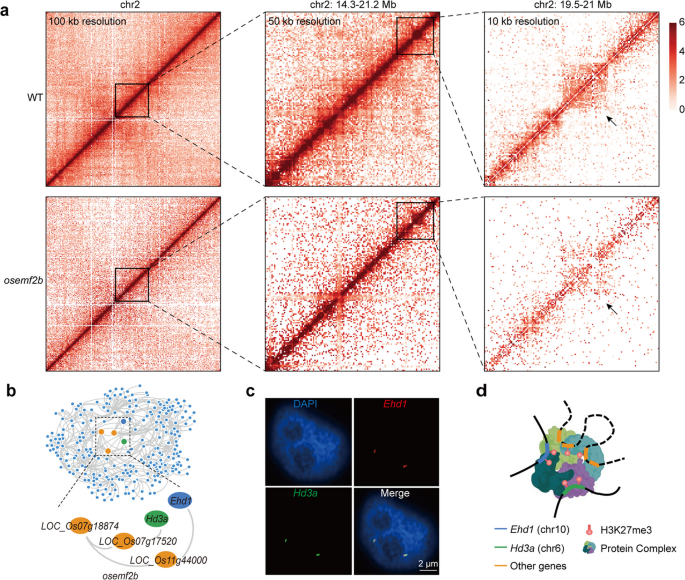
推测:早抽穗1号-抽穗期3a/水稻开花素T1空间簇的形成不依赖于水稻胚胎花2b蛋白,可能存在多梳抑制复合物2其他组分的功能冗余,需进一步敲除其他组分验证。
产品关联:实验所用关键产品:ABclonal的组蛋白H3赖氨酸27三甲基化抗体(货号A2363)。
3.5 高产抽穗期8号相关复合物对早抽穗1号-抽穗期3a/水稻开花素T1簇的结合验证
实验目的:解析反式调控因子高产抽穗期8号、高产抽穗期7号、抽穗期1号对早抽穗1号-抽穗期3a/水稻开花素T1簇的结合机制。
方法细节:构建不同基因型的高产抽穗期8号-绿色荧光蛋白融合近等基因系,包括珍汕97-高产抽穗期7号缺失/高产抽穗期8号-绿色荧光蛋白融合/抽穗期1号功能型、珍汕97-高产抽穗期7号功能型/高产抽穗期8号-绿色荧光蛋白融合/抽穗期1号缺失、珍汕97-高产抽穗期7号功能型/高产抽穗期8号-绿色荧光蛋白融合/抽穗期1号功能型,表型调查开花时间;通过染色质免疫共沉淀测序检测高产抽穗期8号的全基因组结合位点;通过电泳迁移率变动分析验证高产抽穗期8号相关复合物与早抽穗1号、抽穗期3a启动子的体外结合。
结果解读:与野生型相比,珍汕97-高产抽穗期7号缺失/高产抽穗期8号-绿色荧光蛋白融合/抽穗期1号功能型、珍汕97-高产抽穗期7号功能型/高产抽穗期8号-绿色荧光蛋白融合/抽穗期1号缺失、珍汕97-高产抽穗期7号功能型/高产抽穗期8号-绿色荧光蛋白融合/抽穗期1号功能型的开花时间分别延迟11.2天、59.9天、108.1天以上(n=10株,P<0.001);染色质免疫共沉淀测序结果显示高产抽穗期8号在不同近等基因系中均富集在早抽穗1号的启动子区域(翻译起始位点上游-1746至-1082碱基对),以及抽穗期3a的启动子、第三内含子和抽穗期3a与水稻开花素T1的基因间区;电泳迁移率变动分析实验证实高产抽穗期7号/高产抽穗期8号/水稻核因子Y C2亚基和抽穗期1号/高产抽穗期8号/水稻核因子Y C2亚基复合物可结合早抽穗1号启动子的-1374、-1390、-1431位点,以及抽穗期3a启动子的CCAC核心序列,对应结果如下图所示:
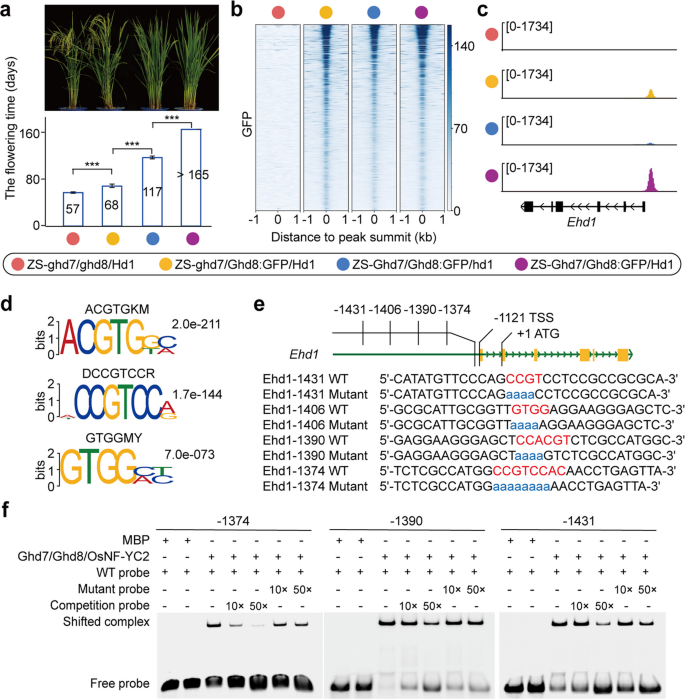
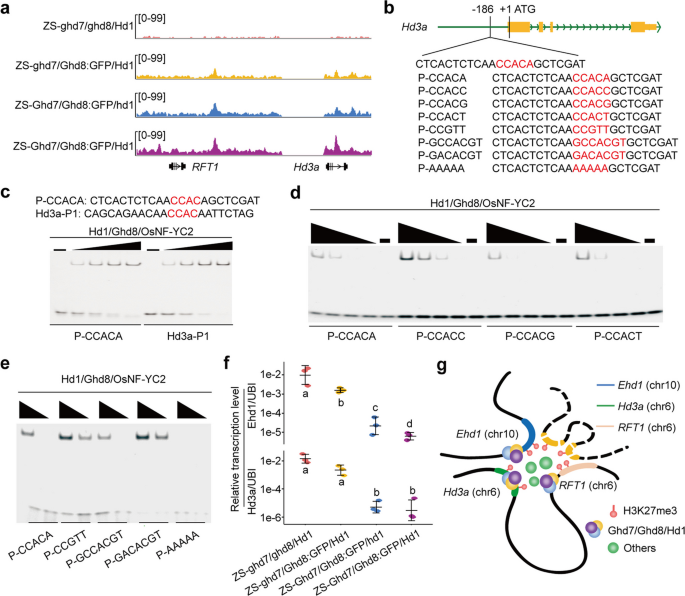
同时,早抽穗1号和抽穗期3a的表达水平在不同近等基因系中呈相似的变化趋势,均与开花时间负相关。
产品关联:实验所用关键产品:Chromotec的绿色荧光蛋白抗体(货号gtma-20)。
3.6 开花调控复合物的液-液相分离特征解析
实验目的:验证高产抽穗期7号、抽穗期1号是否通过液-液相分离形成凝聚体,调控早抽穗1号-抽穗期3a/水稻开花素T1簇的表达。
方法细节:预测抽穗期1号和高产抽穗期7号的内在无序区域;在水稻原生质体中瞬时表达绿色荧光蛋白融合的抽穗期1号和高产抽穗期7号蛋白,通过激光共聚焦显微镜观察凝聚体形成;通过荧光漂白恢复实验检测凝聚体的流动性;在体外表达纯化抽穗期1号和高产抽穗期7号的不同结构域蛋白,验证其液滴形成能力;通过共表达实验验证高产抽穗期8号是否被招募到凝聚体中。
结果解读:抽穗期1号和高产抽穗期7号在水稻原生质体细胞核中形成球形凝聚体,荧光漂白恢复实验显示漂白后的抽穗期1号凝聚体在40秒内恢复约50%的荧光,证实其流动性;体外实验显示抽穗期1号的B盒结构域、CCT结构域和高产抽穗期7号的CCT结构域可形成浓度依赖的液滴,且液滴可发生融合;高产抽穗期8号本身不形成液滴,但可与抽穗期1号或高产抽穗期7号共定位形成凝聚体,对应结果如下图所示:
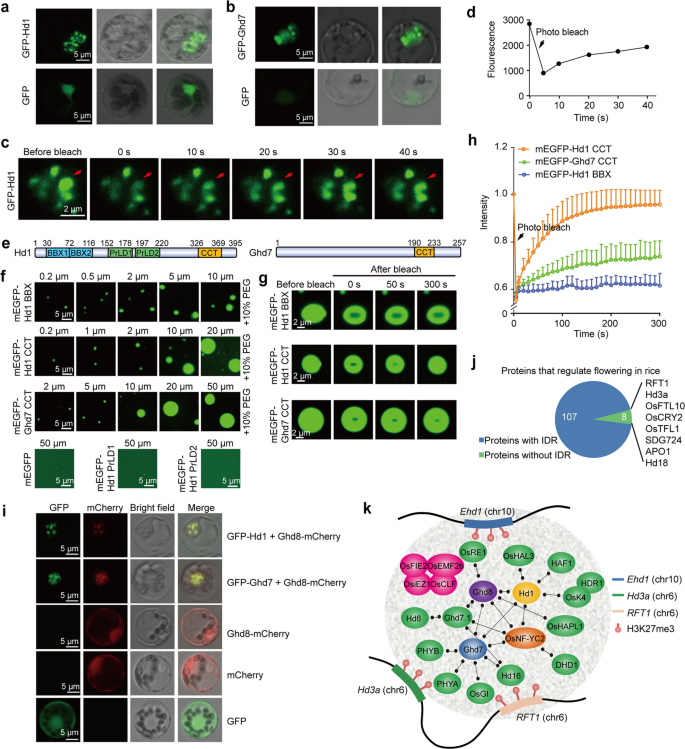
对115个已知水稻开花调控蛋白的预测显示,107个蛋白含有内在无序区域,提示液-液相分离可能是水稻开花调控的普遍机制。
产品关联:文献未提及具体实验产品,领域常规使用蛋白表达纯化试剂、荧光标记抗体和激光共聚焦显微镜。
4. Biomarker研究及发现成果
本研究涉及的生物标志物为组蛋白H3赖氨酸27三甲基化标记的早抽穗1号-抽穗期3a/水稻开花素T1跨染色体抑制型空间簇,筛选逻辑为:通过长读长染色质相互作用配对末端标签测序全基因组筛选互作频率高的基因簇,结合开花调控功能富集,锁定该候选簇;随后通过多组织、多品种、突变体验证其稳定性,最后通过分子生物学实验验证其调控功能。
该生物标志物来源于水稻叶组织细胞核中的染色质,验证方法包括长读长染色质相互作用配对末端标签测序检测染色质互作、杂交链式反应荧光原位杂交验证空间共定位、染色质免疫共沉淀测序验证组蛋白H3赖氨酸27三甲基化修饰、实时荧光定量聚合酶链式反应验证基因表达。特异性与敏感性数据:早抽穗1号和抽穗期3a的杂交链式反应荧光原位杂交共定位率在明恢63中显著高于阴性对照(P<0.05,样本量n=74个细胞核),二者的组蛋白H3赖氨酸27三甲基化水平在四个组织中的相关系数≥0.82,提示该簇的稳定性较高。
核心成果提炼:该空间簇作为水稻开花调控的顺式作用支架,可被高产抽穗期7号/高产抽穗期8号/抽穗期1号液-液相分离凝聚体结合,协同抑制早抽穗1号和抽穗期3a/水稻开花素T1的表达,从而延迟水稻开花;创新性在于首次发现水稻中开花关键基因形成跨染色体的抑制型空间簇,且揭示了液-液相分离介导的反式调控机制;统计学结果显示,高产抽穗期8号近等基因系中,同时具有功能性高产抽穗期7号和抽穗期1号的株系开花时间延迟超过108.1天(n=10,P<0.001),早抽穗1号和抽穗期3a的表达水平显著降低(P<0.01)。该生物标志物可作为水稻开花时间分子育种的表观遗传靶点,通过调控该簇的形成或调控复合物的结合,可精准调节水稻抽穗期,提升品种的区域适应性。