1. 领域背景与文献引入
文献英文标题:Targeting miR-181a/b in retinitis pigmentosa: implications for disease progression and therapy;发表期刊:Cell & Bioscience;影响因子:未公开;研究领域:视网膜退行性疾病(视网膜色素变性)、非编码RNA调控、代谢重编程
视网膜色素变性是一类遗传异质性的退行性疾病,以光感受器进行性死亡为核心特征,最终导致失明,目前已发现超过60个相关致病基因,遗传异质性为疾病机制研究和治疗开发带来巨大挑战。领域共识:视网膜色素变性不仅影响光感受器,还会继发损伤视网膜色素上皮(RPE),而视网膜色素上皮通过营养转运、维持光感受器外节稳态等功能支撑视网膜正常生理,其功能障碍会加速疾病进展。近年来,线粒体功能障碍、氧化应激、糖酵解代谢失调等共同通路成为研究热点,非编码RNA尤其是微小RNA(miRNA)作为基因表达的关键调控因子,被证实参与视网膜色素变性的发病机制,其中miR-181a/b下调已被报道可改善视网膜色素变性小鼠模型的线粒体功能,但该调控在不同视网膜细胞类型中的动态表达模式及具体代谢机制尚未明确,现有研究也未探讨视网膜色素上皮中miR-181a/b的代偿调控机制,这一研究空白限制了miR-181a/b作为治疗靶点的精准应用。本研究针对上述问题,聚焦miR-181a/b在视网膜色素变性进展中神经视网膜与视网膜色素上皮的表达差异,以及视网膜色素上皮特异性下调miR-181a/b的治疗效果与机制,为视网膜色素变性的miRNA靶向治疗提供新的理论依据。
2. 文献综述解析
作者以细胞类型(神经视网膜、视网膜色素上皮)和疾病进展阶段为分类维度,系统梳理了miR-181a/b在代谢调控及视网膜疾病中的研究现状,明确现有研究的局限性并提出本研究的创新方向。
现有研究支持观点方面,已有研究证实miR-181a/b在肿瘤代谢中发挥关键调控作用,比如下调miR-181b-5p可通过上调PDHX抑制胆囊癌细胞的糖酵解过程,过表达miR-181a则通过激活PTEN/AKT通路促进结肠癌细胞的糖酵解增殖;在视网膜疾病领域,前期研究发现广谱下调miR-181a/b可改善视网膜色素变性小鼠的线粒体功能,延缓光感受器死亡,但该研究未区分不同视网膜细胞类型的特异性作用,也未探讨疾病进展中miR-181a/b的动态表达变化。技术方法优势方面,前期研究采用腺相关病毒(AAV)介导的“海绵”载体实现miR-181a/b的广谱下调,可有效改善疾病表型,但该方法无法实现细胞特异性调控,难以解析不同细胞的独立作用;局限性方面,现有研究未关注miR-181a/b在视网膜色素上皮中的代偿机制,也未深入探讨其对视网膜色素上皮代谢重编程的具体影响,缺乏对治疗时效性和细胞特异性的分析。
本研究的创新价值在于,首次揭示了miR-181a/b在视网膜色素变性进展中神经视网膜与视网膜色素上皮的相反表达模式,首次证实视网膜色素上皮中miR-181a/b-1敲除后miR-181a/b-2的代偿调控作用,同时明确了视网膜色素上皮特异性下调miR-181a/b通过抑制有氧糖酵解改善疾病表型的代谢机制,弥补了现有研究在细胞特异性和代谢机制解析上的不足。
3. 研究思路总结与详细解析
本研究的整体目标是明确miR-181a/b在视网膜色素变性进展中的细胞特异性表达模式,探讨视网膜色素上皮特异性下调miR-181a/b的治疗效果及潜在机制,核心科学问题是miR-181a/b在不同视网膜细胞中的动态表达如何调控疾病进展,以及视网膜色素上皮中miR-181a/b的代偿机制与代谢调控路径,技术路线遵循“模型构建→表达分析→表型验证→机制解析”的闭环逻辑,通过细胞特异性基因敲除模型、多维度表型检测及分子机制分析,系统解析miR-181a/b的调控作用。
3.1 实验动物模型构建与验证
实验目的:构建视网膜色素上皮特异性miR-181a/b-1敲除的视网膜色素变性小鼠模型,实现对miR-181a/b的细胞特异性调控,为后续研究提供工具。
方法细节:将携带Pde6βH620Q突变的视网膜色素变性小鼠、Rpe65CreERT2诱导型重组酶小鼠、Mirc14tm1.1Czc(miR-181a/b-1 flox/flox)小鼠进行杂交,经过至少6代回交获得遗传背景一致的实验小鼠,实验组为纯合Pde6βH620Q、纯合miR-181a/b-1 flox/flox且杂合Rpe65CreERT2的小鼠,对照组为纯合Pde6βH620Q、纯合miR-181a/b-1 flox/flox且野生型Rpe65CreERT2的小鼠;在小鼠出生后第8-12天(P8-P12)进行两次腹腔注射他莫昔芬(0.1mg/g体重),诱导视网膜色素上皮中Cre重组酶的激活,从而敲除miR-181a/b-1;通过基因组实时荧光定量PCR(qPCR)验证miR-181a/b-1的敲除效率,同时检测神经视网膜中的敲除情况以确认细胞特异性。
结果解读:基因组PCR结果显示,实验组小鼠视网膜色素上皮中miR-181a/b-1被有效敲除,而神经视网膜中无敲除发生,成功构建了视网膜色素上皮特异性miR-181a/b-1敲除的视网膜色素变性小鼠模型,为后续研究奠定了基础。
实验所用关键产品:他莫昔芬(Sigma-Aldrich)、Phire DNA聚合酶(Fisher Scientific)、TaqMan微小RNA检测试剂盒(Thermo Fisher Scientific)。
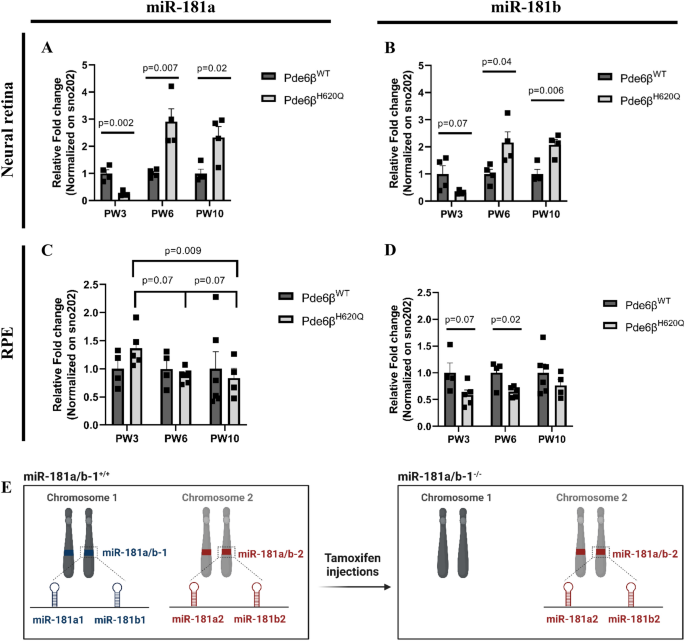
3.2 miR-181a/b在视网膜色素变性进展中的表达模式分析
实验目的:检测视网膜色素变性小鼠疾病进展中神经视网膜与视网膜色素上皮的miR-181a/b表达变化,明确其细胞特异性表达模式。
方法细节:分别在小鼠出生后第3周(PW3,疾病早期)、第6周(PW6,疾病中期)、第10周(PW10,疾病晚期)处死野生型和视网膜色素变性小鼠,分离神经视网膜和视网膜色素上皮组织,提取总RNA,采用TaqMan微小RNA检测试剂盒进行实时荧光定量PCR(qPCR),检测miR-181a和miR-181b的表达水平,以sno202作为内参进行标准化。
结果解读:在神经视网膜中,视网膜色素变性小鼠PW3时miR-181a和miR-181b表达显著下调(n≥4,P<0.05),而PW6和PW10时表达显著上调(n≥4,P<0.05);在视网膜色素上皮中,miR-181b从PW3开始持续下调(n≥4,P<0.05),miR-181a则随疾病进展逐渐下调,PW10时差异显著(n≥4,P<0.05)。这一结果揭示了miR-181a/b在视网膜色素变性进展中的细胞特异性动态表达模式,神经视网膜中miR-181a/b的上调可能促进疾病进展,而视网膜色素上皮中的下调可能是代偿保护反应。
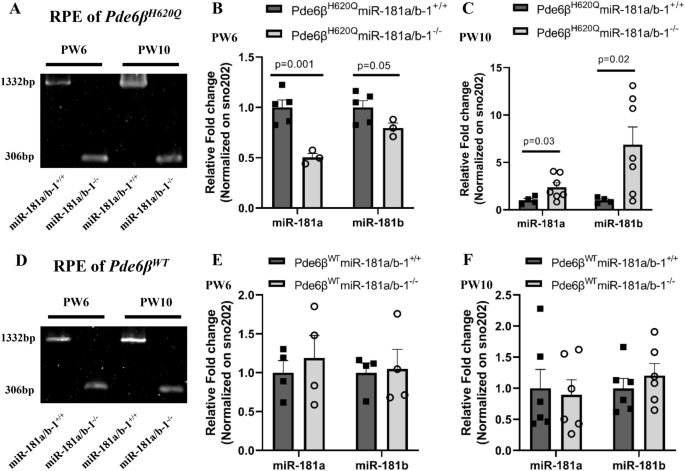
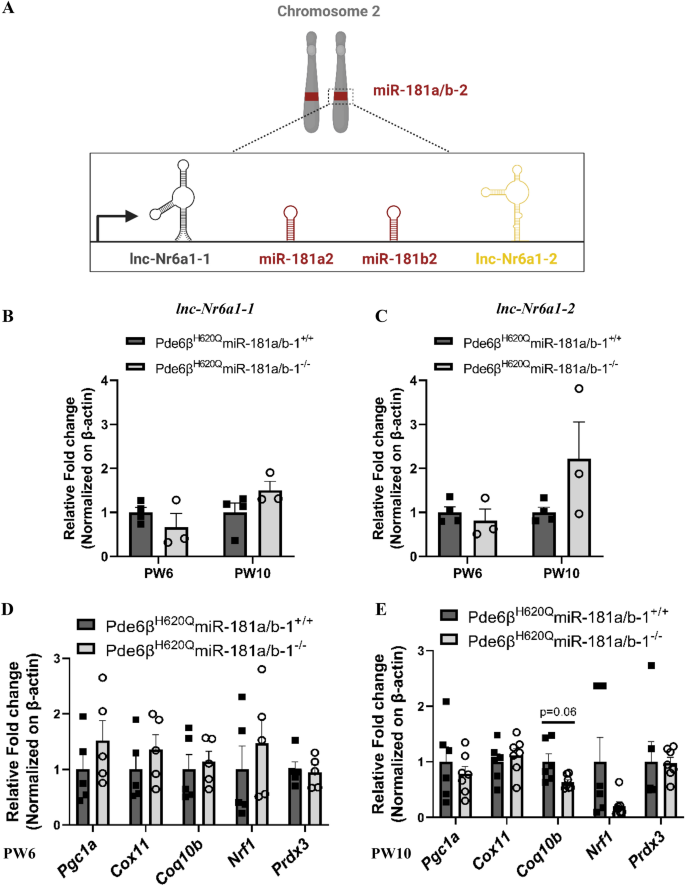
3.3 视网膜色素上皮中miR-181a/b-1敲除后的代偿机制分析
实验目的:探讨视网膜色素上皮特异性敲除miR-181a/b-1后miR-181a/b的表达变化及潜在代偿机制。
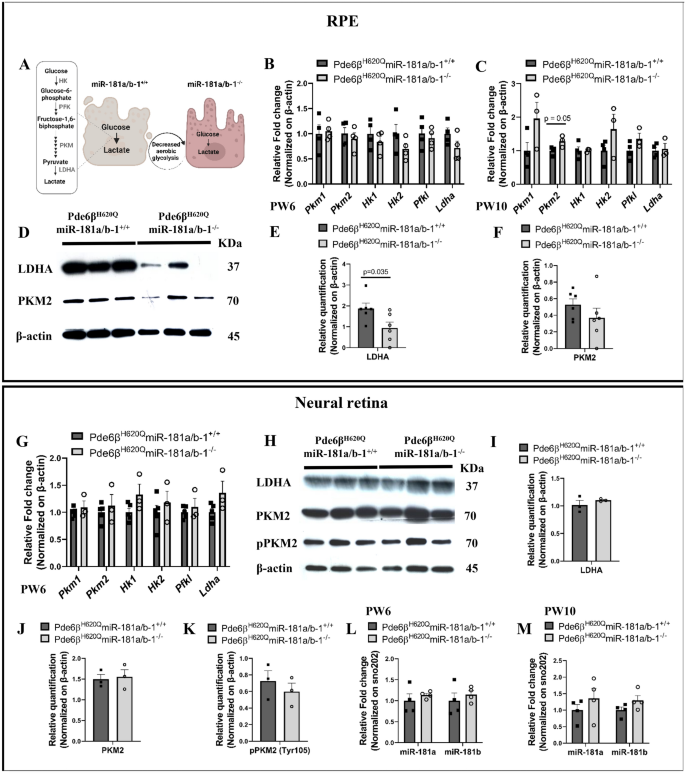
方法细节:分别在PW6和PW10处死实验组和对照组小鼠,分离视网膜色素上皮组织,提取总RNA,采用实时荧光定量PCR(qPCR)检测miR-181a、miR-181b的表达,同时检测miR-181a/b-2的宿主长链非编码RNA lnc-Nr6a1-1和lnc-Nr6a1-2的表达,以及miR-181a/b的线粒体相关靶基因(Nrf1、Cox11、Coq10b等)的表达水平;同时设置野生型背景的视网膜色素上皮特异性miR-181a/b-1敲除小鼠作为对照,排除疾病背景的影响。
结果解读:实验组小鼠PW6时视网膜色素上皮中miR-181a和miR-181b表达显著下调(n≥3,P<0.05),但PW10时表达显著上调(n≥3,P<0.05);而野生型背景的敲除小鼠在两个时间点miR-181a/b表达均无显著变化。实时荧光定量PCR(qPCR)结果显示,PW10时实验组小鼠视网膜色素上皮中lnc-Nr6a1-1和lnc-Nr6a1-2的表达有升高趋势,提示miR-181a/b-2可能代偿miR-181a/b-1的敲除;靶基因表达在PW6时呈升高趋势,PW10时呈降低趋势,与miR-181a/b的表达变化负相关,进一步支持代偿机制的存在。这一结果首次揭示了视网膜色素上皮中miR-181a/b家族的代偿调控机制,为miRNA靶向治疗的时效性提供了理论依据。
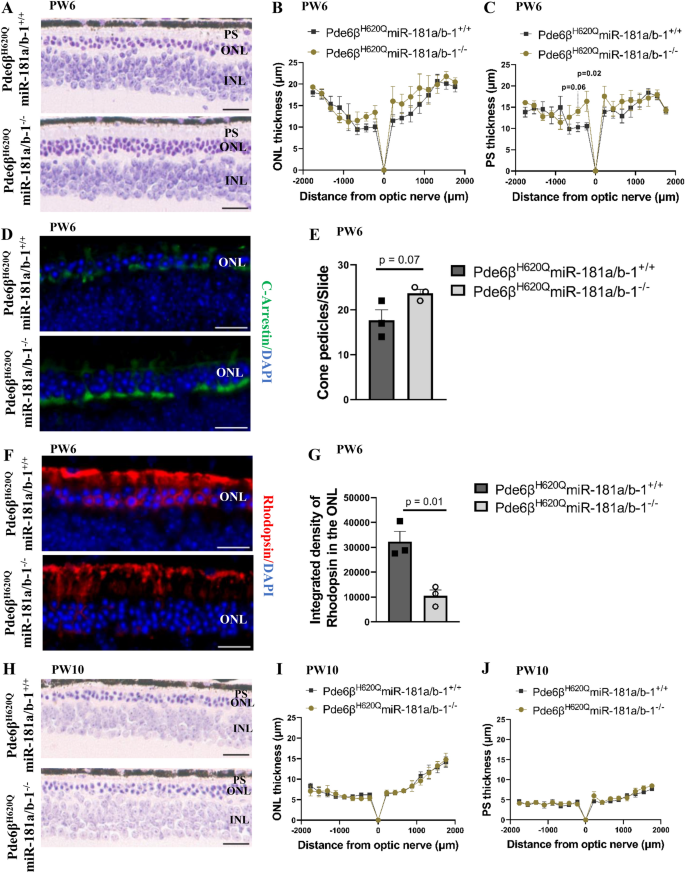
3.4 视网膜色素上皮特异性下调miR-181a/b对视网膜色素变性表型的影响
实验目的:评估视网膜色素上皮中miR-181a/b下调对视网膜色素变性小鼠视网膜变性的治疗效果,明确其时效性。
方法细节:分别在PW6和PW10对实验组和对照组小鼠进行表型检测,包括组织学分析、视网膜电图(ERG)检测、免疫荧光染色。组织学分析采用苏木精-伊红(H&E)染色,测量视网膜外核层(ONL)厚度和光感受器外节长度;ERG检测评估视网膜的光反应功能;免疫荧光染色采用视紫红质抗体标记视杆细胞,C-Arrestin抗体标记视锥细胞,分析光感受器的形态和数量。
结果解读:PW6时,实验组小鼠视网膜外核层厚度有增加趋势,光感受器外节长度显著延长(n≥3,P<0.05);免疫荧光结果显示,实验组小鼠视锥细胞足突数量显著增加(n=3,P<0.05),视紫红质在视杆细胞外节的定位更正常,提示光感受器变性被暂时延缓;但PW10时,这些形态学改善消失,实验组与对照组无显著差异;ERG检测在两个时间点均未检测到显著的功能差异。这一结果表明,视网膜色素上皮中miR-181a/b下调可暂时延缓视网膜色素变性的进展,但由于miR-181a/b-2的代偿作用,治疗效果无法持续。
3.5 视网膜色素上皮中miR-181a/b下调的代谢机制分析
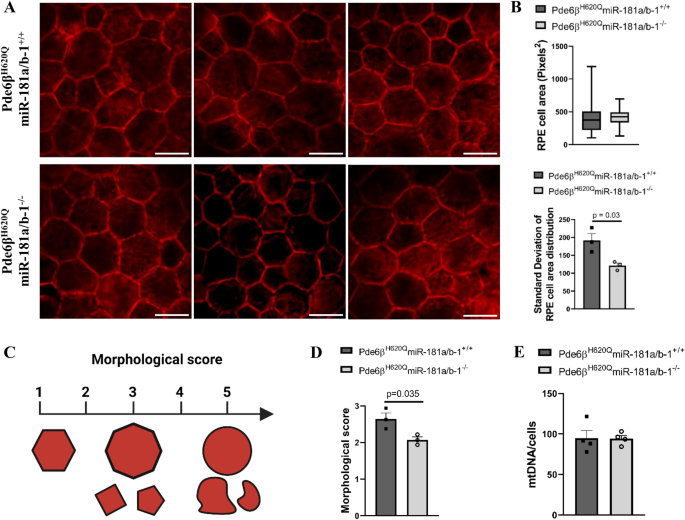
实验目的:探讨视网膜色素上皮中miR-181a/b下调对视网膜色素上皮形态、线粒体功能及糖酵解代谢的影响,明确其治疗机制。
方法细节:PW6时分离实验组和对照组小鼠的视网膜色素上皮铺片,采用鬼笔环肽染色分析视网膜色素上皮细胞的形态,通过盲法评分评估细胞健康状态;采用实时荧光定量PCR(qPCR)检测线粒体DNA(mtDNA)含量,评估线粒体生物发生;采用蛋白质免疫印迹(Western blot)检测糖酵解相关蛋白乳酸脱氢酶A(LDHA)、丙酮酸激酶M2(PKM2)的表达水平。
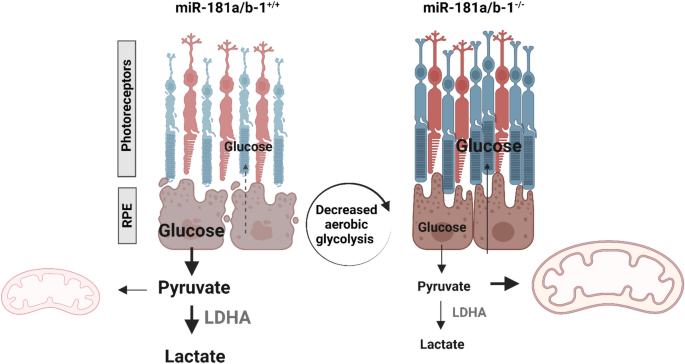
结果解读:实验组小鼠视网膜色素上皮细胞形态更均匀,细胞大小和形状的异质性更低,盲法健康评分显著高于对照组(n=3,P<0.05);线粒体DNA含量无显著变化,提示miR-181a/b下调未增加线粒体数量,但可能改善线粒体功能;蛋白质免疫印迹结果显示,实验组小鼠视网膜色素上皮中LDHA蛋白水平显著降低(n=6,P<0.05),而PKM2蛋白水平无显著变化,提示miR-181a/b下调抑制了视网膜色素上皮的有氧糖酵解代谢。这一结果表明,视网膜色素上皮中miR-181a/b下调通过抑制有氧糖酵解,改善视网膜色素上皮形态,从而暂时延缓光感受器变性。
4. Biomarker研究及发现成果
本研究中涉及的生物标志物为miR-181a/b,属于组织特异性非编码RNA生物标志物,其在视网膜色素变性进展中的细胞特异性表达模式可作为疾病进展的评估指标,同时也是潜在的治疗靶点。
Biomarker定位:miR-181a/b作为视网膜色素变性的细胞特异性生物标志物,筛选逻辑为在视网膜色素变性小鼠模型中动态检测疾病进展不同阶段神经视网膜与视网膜色素上皮的表达变化,验证逻辑为通过细胞特异性敲除模型验证其对疾病表型的调控作用及机制。
研究过程详述:miR-181a/b的来源为小鼠神经视网膜和视网膜色素上皮组织,验证方法包括实时荧光定量PCR(qPCR)检测表达水平、免疫荧光染色评估表型变化、蛋白质免疫印迹检测下游代谢蛋白表达;特异性方面,神经视网膜中miR-181a/b在疾病中期和晚期显著上调,而视网膜色素上皮中miR-181b从疾病早期开始持续下调,miR-181a随疾病进展逐渐下调,呈现细胞特异性的表达模式;敏感性方面,在疾病早期(PW3)即可检测到视网膜色素上皮中miR-181b的显著下调(n≥4,P<0.05),可作为疾病早期的分子信号。
核心成果提炼:miR-181a/b可作为视网膜色素变性进展的细胞特异性生物标志物,神经视网膜中miR-181a/b的上调提示疾病进展,视网膜色素上皮中的下调提示代偿保护反应;同时,视网膜色素上皮特异性下调miR-181a/b可通过抑制有氧糖酵解改善视网膜色素上皮形态,暂时延缓光感受器变性,首次揭示了miR-181a/b家族在视网膜色素上皮中的代偿调控机制,为视网膜色素变性的miRNA靶向治疗提供了新的靶点和策略,具有重要的转化应用价值。