1. 领域背景与文献引入
文献英文标题:The histone methyltransferase SDG8 mediates the epigenetic modification of light and carbon responsive genes in plants;发表期刊:Genome Biology;影响因子:未公开;研究领域:植物表观遗传学(组蛋白甲基化调控环境响应基因机制)
表观遗传调控是植物响应环境刺激(如光、碳源)和发育信号的核心机制之一,组蛋白共价修饰作为表观遗传的重要组成部分,不同赖氨酸位点的甲基化对应不同的转录状态,比如组蛋白H3赖氨酸36三甲基化(H3K36me3)是公认的转录激活标记,而组蛋白H3赖氨酸9二甲基化(H3K9me2)则与基因沉默相关。在酵母和动物中,H3K36甲基转移酶(如酵母SET2)的功能已被广泛研究,但其在植物中的调控机制仍不清晰。植物中SET结构域组(SDG)家族包含32个成员,其中SDG8的研究多集中在单基因层面,发现其参与开花时间调控、防御反应、色素合成等过程,但缺乏全基因组水平的组蛋白修饰与转录组整合分析,且SDG8如何调控光和碳响应基因的表观机制仍未明确。本研究以拟南芥sdg8-5缺失突变体为材料,通过多组学技术系统解析SDG8的全基因组组蛋白甲基化功能,填补了植物表观遗传调控环境信号响应机制的研究空白,为理解植物能量代谢的表观调控网络提供了新的视角。
2. 文献综述解析
作者围绕植物表观遗传调控的核心作用、组蛋白甲基化的功能分类、SDG家族的研究现状及SDG8的已知功能展开综述,逻辑清晰地从表观遗传的整体作用过渡到SDG8的具体研究缺口。现有研究已证实组蛋白甲基化在植物发育和环境响应中的关键作用,不同赖氨酸位点的甲基化对应不同的转录状态;在SDG家族研究中,酵母和动物的H3K36甲基转移酶功能已明确,而植物中SDG8的研究多集中在单基因层面,初步建立了其与开花、防御等表型的关联,但这些研究仅揭示了SDG8的部分功能,缺乏全基因组水平的组蛋白甲基化分析,也未明确SDG8在光和碳信号响应中的表观调控机制。现有研究的优势在于为SDG8的功能研究奠定了基础,局限性在于未整合表观组和转录组数据解析其全基因组调控网络,且对SDG8靶基因的特异性调控机制了解不足。本研究的创新价值在于首次在全基因组层面解析了SDG8对组蛋白H3赖氨酸36三甲基化的调控模式,结合染色质免疫共沉淀测序(ChIP-Seq)和转录组数据鉴定了SDG8的直接靶基因,明确了其在光和碳响应基因调控中的核心作用,填补了植物表观遗传调控环境信号响应机制的研究空白。
3. 研究思路总结与详细解析
本研究以拟南芥sdg8-5缺失突变体为核心材料,整体研究思路遵循“突变体鉴定→表观组分析→靶基因鉴定→转录组整合→功能验证”的闭环逻辑,旨在明确SDG8的全基因组组蛋白甲基化功能及对光和碳响应基因的调控机制,核心科学问题是SDG8如何通过组蛋白H3赖氨酸36三甲基化修饰特异性调控靶基因的表达,进而参与植物对光和碳信号的响应。
3.1 突变体鉴定与互补验证
实验目的是定位cli186突变体的致病基因,并通过互补实验验证SDG8的功能。方法细节:采用Affymetrix ATH1芯片对cli186突变体和野生型的基因组DNA进行杂交,定位缺失区域,随后通过PCR扩增验证缺失的具体范围;构建包含SDG8完整基因组序列(含启动子、内含子)的载体,通过农杆菌介导的花序浸染法转化cli186突变体,获得互补株系后,检测开花时间、ASN1基因表达等表型。结果解读:芯片分析显示cli186突变体在1号染色体上存在13.8 kb的缺失,包含SDG8的完整编码区;互补株系恢复了野生型的开花时间和ASN1基因的光/碳响应表达模式,证实SDG8是cli186突变体的致病基因,将其重命名为sdg8-5。产品关联:文献未提及具体实验产品,领域常规使用植物基因组DNA提取试剂、PCR扩增试剂、农杆菌转化载体、基因芯片等。
3.2 全基因组组蛋白甲基化谱分析
实验目的是解析sdg8-5突变体中组蛋白甲基化的全基因组变化,明确SDG8的组蛋白甲基化特异性。方法细节:选取3周龄光下生长的野生型和sdg8-5突变体植株,进行2小时的碳和光处理后,分别用抗组蛋白H3赖氨酸36三甲基化抗体(Abcam)和抗组蛋白H3赖氨酸4三甲基化抗体(Upstate/Millipore)进行染色质免疫共沉淀测序,每个处理设置2个生物学重复;采用SICER软件分析两组间的差异组蛋白修饰区域。结果解读:染色质免疫共沉淀测序结果显示,sdg8-5突变体中有4060个基因的组蛋白H3赖氨酸36三甲基化修饰水平显著降低(FDR<0.05,倍数变化>2),而组蛋白H3赖氨酸4三甲基化修饰水平几乎无差异;进一步分析发现,组蛋白H3赖氨酸36三甲基化的降低主要集中在基因的3"端区域,与酵母中SET2的作用模式类似,提示SDG8参与转录延伸过程。
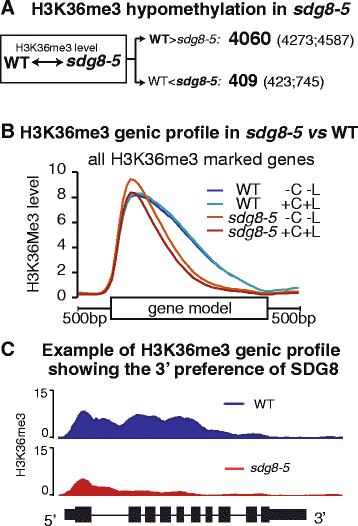
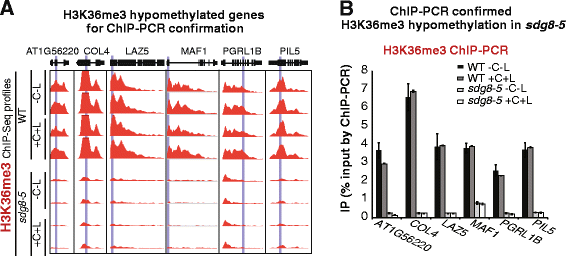
随后通过染色质免疫共沉淀-PCR验证了6个代表性基因的组蛋白H3赖氨酸36三甲基化低甲基化,与染色质免疫共沉淀测序结果一致。
产品关联:使用了抗组蛋白H3赖氨酸36三甲基化抗体(Abcam)、抗组蛋白H3赖氨酸4三甲基化抗体(Upstate/Millipore)、染色质免疫共沉淀试剂盒、Illumina测序平台及相关文库构建试剂。
3.3 SDG8直接靶基因的鉴定
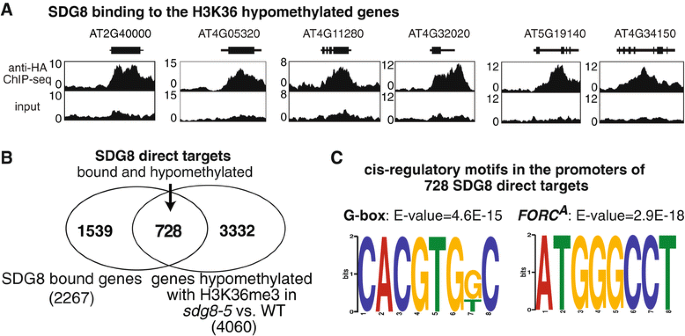
实验目的是筛选SDG8直接结合的靶基因,明确其特异性调控的基因集合。方法细节:构建C端带有HA标签的SDG8互补载体,转化sdg8-5突变体获得功能互补的转基因株系(hSDG8);对2.5周龄的hSDG8植株进行抗HA抗体的染色质免疫共沉淀测序,分析SDG8的全基因组结合区域。结果解读:染色质免疫共沉淀测序共鉴定到2267个SDG8直接结合的基因,其中728个基因同时在sdg8-5突变体中表现为组蛋白H3赖氨酸36三甲基化低甲基化,这些直接靶基因显著富集在防御反应、光合作用、营养代谢和能量代谢等生物学过程中。
产品关联:使用了抗HA抗体(Abcam)、染色质免疫共沉淀测序相关试剂及测序平台。
3.4 转录组分析与多组学数据整合
实验目的是分析sdg8-5突变体的基因表达变化,整合表观组与转录组数据揭示组蛋白H3赖氨酸36三甲基化与基因表达的关联。方法细节:对3周龄处理后的野生型和sdg8-5植株进行RNA提取,采用Affymetrix ATH1基因芯片进行转录组分析,设置3个生物学重复;通过三因素ANOVA分析基因型、光处理、碳处理及交互作用对基因表达的影响,随后整合染色质免疫共沉淀测序的组蛋白H3赖氨酸36三甲基化数据与转录组数据。结果解读:转录组分析显示,sdg8-5突变体中有2158个基因表达显著下调,1923个基因表达显著上调;整合分析发现,下调基因中有1084个基因同时表现为组蛋白H3赖氨酸36三甲基化低甲基化(P<1e-239),说明组蛋白H3赖氨酸36三甲基化修饰与基因表达水平呈正相关,SDG8通过维持组蛋白H3赖氨酸36三甲基化水平促进靶基因的高表达。产品关联:使用了RNeasy植物RNA提取试剂盒(Qiagen)、Affymetrix ATH1基因芯片及相关杂交试剂。
3.5 功能验证与调控机制分析
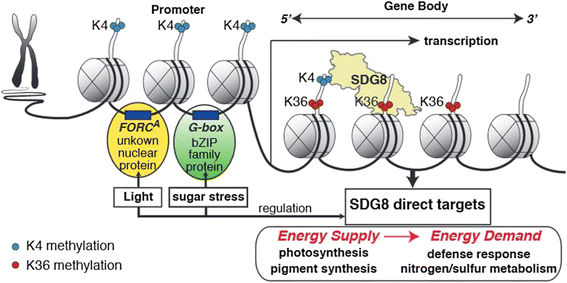
实验目的是验证SDG8对光合作用的调控作用,并解析其靶基因的特异性调控机制。方法细节:测定不同光强下生长的sdg8-5突变体和野生型植株的叶绿素含量;采用MEME软件分析728个SDG8直接靶基因的启动子区域,筛选富集的顺式作用元件。结果解读:sdg8-5突变体的叶绿素含量显著低于野生型(n=4,P<0.05),证实SDG8调控光合作用相关基因的表达;启动子基序分析显示,靶基因启动子显著富集G-box(bZIP转录因子结合基序)和FORC^A基序,提示SDG8可能与bZIP转录因子协同作用,特异性调控靶基因的组蛋白H3赖氨酸36三甲基化修饰及表达。
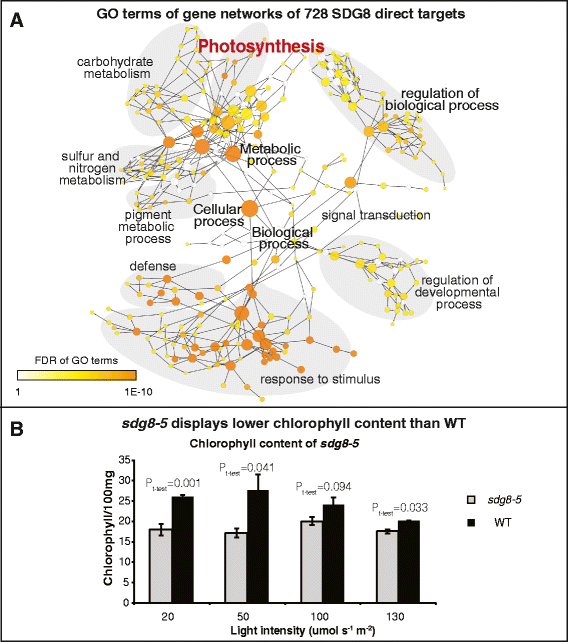
产品关联:文献未提及具体实验产品,领域常规使用叶绿素提取试剂、荧光分光光度计、MEME在线分析工具等。
4. Biomarker研究及发现成果
本研究中的核心Biomarker为组蛋白H3赖氨酸36三甲基化修饰标记,以及SDG8调控的光和碳响应靶基因,通过全基因组筛选与验证,明确了其在植物环境响应中的功能与调控机制。
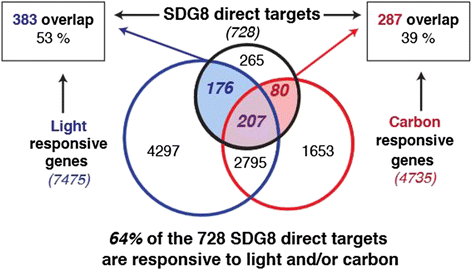
组蛋白H3赖氨酸36三甲基化作为转录激活型组蛋白修饰标记,其筛选逻辑为通过染色质免疫共沉淀测序比较野生型与sdg8-5突变体的组蛋白修饰差异,结合SDG8的染色质免疫共沉淀测序数据,鉴定出SDG8直接调控的靶基因上的组蛋白H3赖氨酸36三甲基化标记;SDG8靶基因作为功能Biomarker,筛选逻辑为同时满足SDG8直接结合且组蛋白H3赖氨酸36三甲基化修饰依赖SDG8的基因集合。研究过程详述:组蛋白H3赖氨酸36三甲基化标记的来源为拟南芥染色质样本,验证方法为染色质免疫共沉淀测序和染色质免疫共沉淀-PCR,特异性表现为SDG8调控的组蛋白H3赖氨酸36三甲基化主要富集在基因的3"端区域,敏感性为在4060个基因中检测到组蛋白H3赖氨酸36三甲基化低甲基化,其中728个为SDG8直接靶基因;SDG8靶基因的验证方法为染色质免疫共沉淀测序和转录组分析,其中64%的直接靶基因响应光和/或碳信号(P<4.2E-17),提示这些基因在植物环境响应中的核心作用。
核心成果提炼:组蛋白H3赖氨酸36三甲基化标记与SDG8靶基因的高表达水平正相关,SDG8通过维持这些靶基因的组蛋白H3赖氨酸36三甲基化修饰,调控光合作用、营养代谢和能量代谢等生物学过程;SDG8直接靶基因中富集的G-box基序提示其与bZIP转录因子协同调控光和碳响应基因的表达,为植物表观遗传调控环境信号响应的机制提供了新的模型。此外,研究还发现SDG8参与介导光和碳信号诱导的组蛋白H3赖氨酸36三甲基化修饰变化,野生型中有54个基因在处理后组蛋白H3赖氨酸36三甲基化水平升高,而sdg8-5突变体中仅9个基因有此变化,进一步证实SDG8在环境信号的表观调控中的关键作用。