1. 领域背景与文献引入
文献英文标题:EphA5 and EphA6: regulation of neuronal and spine morphology;发表期刊:Cell Bioscience;影响因子:未公开;研究领域:神经科学(大脑皮层发育与神经元形态调控)
领域共识:Eph/ephrin家族的双向信号通路是神经发育中神经元迁移、轴突导向及突触可塑性的核心调控机制,通过形成分子梯度介导细胞的吸引或排斥反应。
Eph家族受体酪氨酸激酶是脊椎动物基因组中最大的相关激酶家族,包含16个成员,分为EphA和EphB两个亚类,通过与ephrin配体的相互作用,在胚胎发育及出生后组织功能中发挥关键作用,涵盖组织分区、血管生成、轴突导向及突触可塑性等多个生物学过程。大脑皮层作为调控高级认知行为、运动控制及情绪反应的核心脑区,其精确发育依赖严格的分子调控网络,Eph/ephrin家族成员在皮层高表达,是皮层分区和神经元定位的核心调控分子。
现有研究已证实EphA5和EphA6在大脑皮层的特定层中高表达,且功能缺失会导致一系列行为学异常:EphA5敲除小鼠出现空间导航障碍、攻击行为减弱,EphA6敲除小鼠表现出学习记忆缺陷,但这些行为学异常对应的神经元形态学改变尚未被系统解析,领域内存在行为学表型与形态学机制的关联空白。本研究以EphA5、EphA6单敲及双敲小鼠为模型,通过Golgi染色、免疫组化等技术,系统解析这两个受体对皮层神经元聚集、树突及树突棘形态的调控作用,填补了神经发育领域中Eph受体功能与神经元结构关联的研究空白,为理解神经发育障碍的机制提供了形态学依据。
2. 文献综述解析
作者围绕Eph家族在神经发育中的调控作用,重点梳理了EphA5和EphA6的功能研究现状,明确了行为学缺陷与神经元形态改变的关联缺失是核心研究空白,为本次研究的开展奠定了逻辑基础。
现有研究中,Eph家族被划分为EphA和EphB两个亚类,多数EphA受体与GPI锚定的ephrin-A配体结合,EphB受体与跨膜的ephrin-B配体结合,同时存在部分交叉结合的情况,如EphA4可结合ephrin-B配体,EphB2对ephrin-A5有亲和力。在大脑皮层发育过程中,Eph/ephrin家族成员的表达梯度形成了关键的位置信号,指导神经元迁移至正确的皮层层,并参与突触的形成与可塑性调控。已有研究主要聚焦于EphA5和EphA6的行为学功能,通过基因敲除或显性负性突变模型,发现其与空间导航、学习记忆、攻击行为等高级神经功能相关,但这些研究未深入解析行为学异常背后的神经元结构变化,缺乏形态学层面的直接证据,尤其是树突棘形态这一突触功能的关键结构基础的分析。
本研究的创新价值在于,首次将EphA5和EphA6的功能研究从行为学层面拓展至形态学层面,通过系统的Golgi染色和免疫组化分析,明确了这两个受体对皮层神经元分布和树突棘形态的调控作用,建立了行为学缺陷与神经元结构异常的直接关联,弥补了现有研究中形态学机制缺失的不足,为Eph家族在神经发育中的功能研究提供了新的视角。
3. 研究思路总结与详细解析
本研究以“EphA5和EphA6对皮层神经元形态的调控作用”为核心目标,聚焦“Eph受体缺失如何导致神经元结构异常”这一科学问题,采用“基因敲除模型构建→表达模式分析→形态学检测→细胞类型验证”的闭环技术路线,系统解析了两个受体在皮层神经元聚集、树突棘发育中的功能。
3.1 基因敲除小鼠模型构建与验证
本环节的实验目的是构建EphA5、EphA6单敲及双敲小鼠模型,为后续形态学分析提供稳定的实验材料。方法细节上,EphA5 LacZ/LacZ小鼠采用实验室已报道的构建方法,EphA6敲除小鼠通过同源重组技术,将LacZ编码序列插入EphA6基因的外显子3,筛选获得纯合敲除的EphA6 LacZ/LacZ小鼠;随后将EphA5和EphA6单敲小鼠杂交,获得双敲小鼠,所有小鼠均饲养于符合Rutgers大学实验动物护理与使用指南的标准环境中。结果解读显示,成功构建三种基因敲除模型,双敲小鼠无明显生理和生殖异常,可用于后续实验分析。实验所用关键产品:文献未提及具体实验产品,领域常规使用基因编辑试剂盒、PCR引物、动物饲养设备等试剂/仪器。
3.2 EphA5和EphA6在成年小鼠脑内的表达检测
本环节的实验目的是明确EphA5和EphA6在成年小鼠脑内的表达分布,为形态学分析提供组织学基础。方法细节上,采用β-半乳糖苷酶染色法,对2月龄杂合和纯合敲除小鼠的脑切片进行染色,检测LacZ标记的受体表达情况。结果解读显示,EphA5在大脑皮层、杏仁核、梨状皮层、海马高表达,皮层II/III、IV、V层信号更为显著,丘脑、纹状体仅有微弱表达;EphA6在眶皮层、嗅球、皮层II/III/V层等广泛脑区高表达,两者在皮层的表达区域高度重叠,提示它们在皮层神经元调控中可能存在协同或互补作用(n=3)。
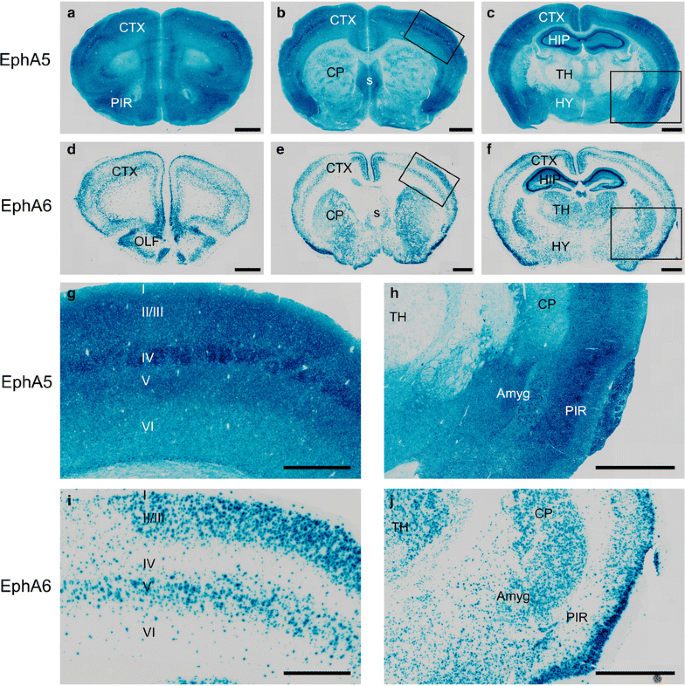
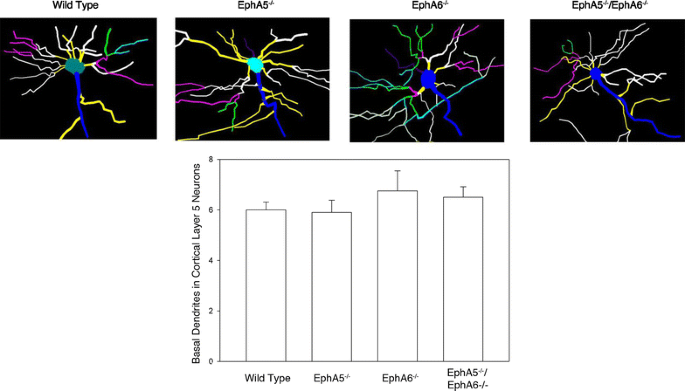
3.3 皮层神经元聚集形态分析
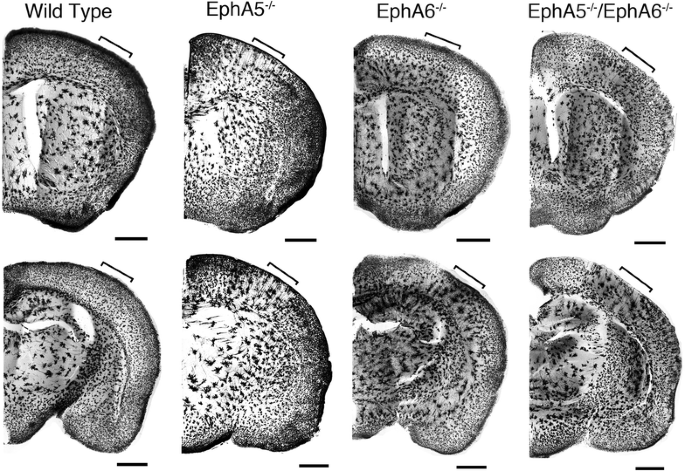
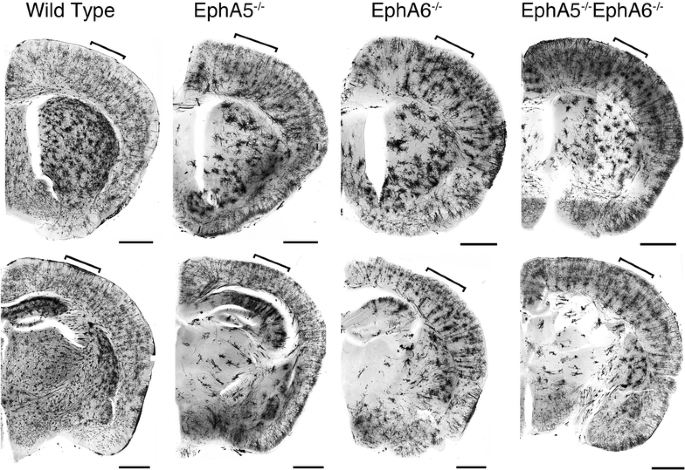
本环节的实验目的是检测EphA5和EphA6缺失对皮层神经元分布的影响。方法细节上,采用两种Golgi染色方案:第一种是对经4%多聚甲醛灌注的小鼠脑组织进行染色,第二种是对新鲜未灌注的脑组织直接染色,对2月龄小鼠的脑切片进行染色后观察皮层神经元的分布情况。结果解读显示,两种染色方案均发现,EphA5、EphA6单敲及双敲小鼠的大脑皮层出现大量异常的神经元聚集,而野生型小鼠的神经元分布均匀;其中EphA6敲除小鼠的神经元聚集现象更为显著,双敲小鼠的表型与EphA6单敲小鼠相似,提示EphA6在调控皮层神经元分布中起主导作用;新鲜脑组织染色结果显示聚集主要为神经元,而灌注染色显示聚集包含神经元和胶质细胞(n=3)。
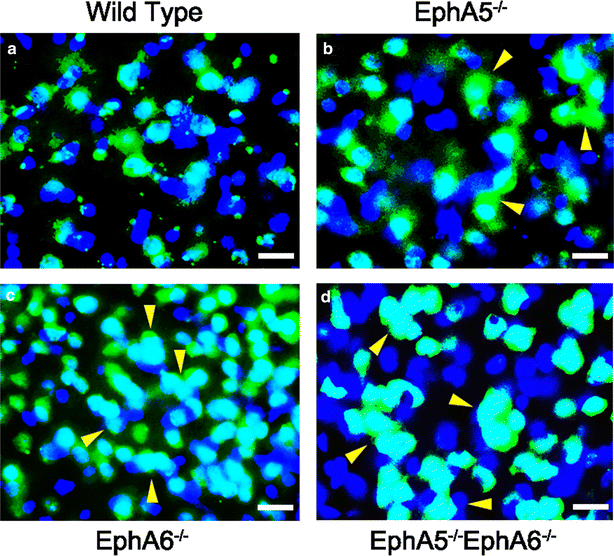
3.4 神经元聚集的细胞类型验证
本环节的实验目的是确认异常聚集细胞的类型,明确是否包含神经元和非神经元细胞。方法细节上,采用免疫组化技术,用神经元特异性标记物NeuN和细胞核标记物DAPI对小鼠脑切片进行双重染色,观察聚集区域的细胞类型。结果解读显示,聚集区域同时存在NeuN阳性的神经元和NeuN阴性的非神经元细胞,证实神经元聚集现象涉及神经元和胶质细胞两类细胞,进一步支持了Golgi染色的结果(n=3)。
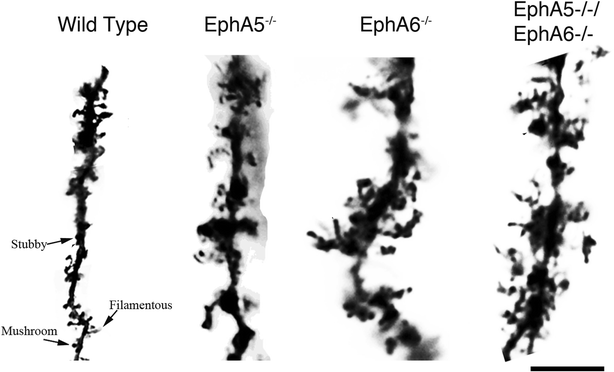
3.5 皮层V层神经元基底树突与树突棘形态分析
本环节的实验目的是检测EphA5和EphA6缺失对皮层神经元树突及树突棘形态的影响。方法细节上,采用Neurolucida软件绘制皮层V层神经元的基底树突,统计树突数量;通过Golgi染色观察树突棘的形态特征。结果解读显示,统计分析60个神经元(每基因型3只小鼠,每只20个神经元)后发现,三种敲除模型与野生型小鼠的基底树突数量无显著差异(P>0.05);但树突棘形态出现显著异常,野生型小鼠的树突棘形态清晰,可分为丝状、短粗、蘑菇状三类,而敲除小鼠的树突棘无法按常规标准分类,形成类似花朵的重叠结构,树突棘颈部与头部边界模糊,提示树突棘的成熟过程受到严重影响。
4. Biomarker研究及发现成果
本研究未涉及疾病诊断或预后类生物标志物(Biomarker),而是鉴定了EphA5和EphA6作为调控皮层神经元形态的关键功能分子标志物,解析了其在神经元聚集和树突棘发育中的核心作用。
Biomarker定位方面,EphA5和EphA6属于Eph家族受体酪氨酸激酶,作为神经元形态调控的功能标志物,其筛选逻辑基于前期研究中两者在皮层的高表达及行为学功能提示,通过基因敲除模型验证其对神经元形态的调控作用。研究过程详述显示,该Biomarker的来源为小鼠脑皮层组织,验证方法包括基因敲除模型构建、Golgi染色、免疫组化技术,特异性表现为在皮层II/III/V层高表达,功能验证显示其缺失会导致神经元聚集和树突棘形态异常,由于属于功能调控标志物而非疾病诊断标志物,无敏感性和特异性的ROC曲线数据。核心成果提炼显示,本研究首次发现EphA5和EphA6调控皮层神经元的分布和树突棘形态,其中EphA6发挥主导作用,其功能缺失导致的树突棘异常可能是行为学缺陷的形态学基础,创新性在于建立了Eph受体功能与神经元形态的直接关联,为神经发育障碍的机制研究提供了新的视角;统计数据方面,仅基底树突数量统计显示P>0.05,其余形态学结果为定性描述,无具体统计学P值。